你的免疫系统几岁了?它比你的日历年龄更重要!最新综述深度解读“免疫衰老时钟”:利用基因组、转录组、蛋白组等多组学数据,结合AI模型,不仅精准估算免疫年龄,更能预测疾病风险。本文构建“任务-模态-模型”框架,系统梳理了从数据到临床应用的完整流程,揭示免疫时钟如何从科研走向精准健康管理。
今天给大家解读一篇2月发表在《Cells》上的题目为“Immune Ageing Clocks: A Methods-Oriented Review of Tasks, Modalities, Models, and Recalibration.”的文章。本综述旨在解决免疫衰老时钟研究因任务定义、检测方案和评估标准异质而导致的局限。文章首先介绍了免疫衰老的背景及构建整合性生物标志物的必要性。随后,提出了核心的“任务-模态-模型”框架,并详细阐述了构建免疫衰老时钟的端到端方法,涵盖核心数据类型、建模方法和工作流程标准化。接着,应用该框架对现有研究进行了分类评述(年龄时钟、风险时钟、细胞谱系时钟)。然后,系统分析了当前研究在方法学、生物学和证据标准方面面临的挑战。最后,展望了未来发展方向,包括多模态整合、组织与细胞类型特异性时钟、干预导向的时钟以及建立可共享的“免疫年龄”度量语言。(请持续关注我们,每天为您解读最新见刊的文献!)想薅生信资料羊毛?直接在对话框回复 “资料”,免费领取干货大礼包!
不想做实验,没数据,还想要快速发表文章,没问题的!公共数据库就是我们的数据宝藏!没思路不用担心,作为专业的生信团队,我们很乐意为你们效劳,提供研究路线设计和数据挖掘分析,扫码联系我们吧!


团队成员合影(位于上海陆家嘴中心,可随时预约参观)
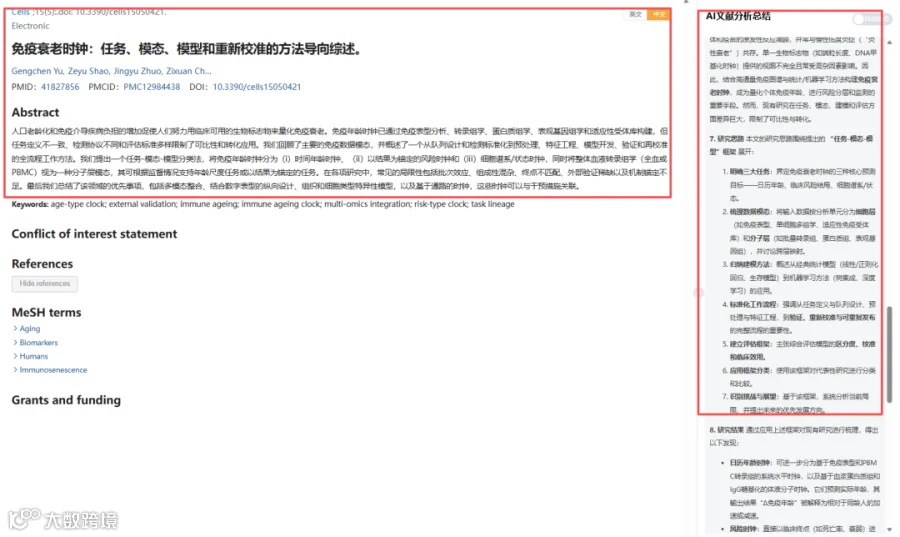
题目:《免疫衰老时钟:任务、模态、模型和重新校准的方法导向综述》Immune Ageing Clocks: A Methods-Oriented Review of Tasks, Modalities, Models, and Recalibration
发表期刊:Cells
影响因子:5.2
研究背景:
人口快速老龄化对全球公共卫生构成挑战。年龄是慢性进展性疾病的主要风险因素。在机制层面,免疫衰老是指与年龄相关的免疫系统重塑,其特征包括对病原体和疫苗的原发性反应减弱,并常与慢性低度炎症(“炎性衰老”)共存。单一生物标志物(如端粒长度、DNA甲基化时钟)提供的视图不完全且常受混杂因素影响。因此,结合高通量免疫图谱与统计/机器学习方法构建免疫衰老时钟,成为量化个体免疫年龄、进行风险分层和监测的重要手段。然而,现有研究在任务、模态、建模和评估方面差异巨大,限制了可比性与转化。
CNSknowall 平台 Pubmed+AI 快速提炼全文要点
研究思路:
本文的研究思路围绕提出的 “任务-模态-模型”框架 展开:
- 明确三大任务
界定免疫衰老时钟的三种核心预测目标——日历年龄、临床风险结局、细胞谱系/状态。
- 梳理数据模态
将输入数据按分析单元分为细胞层(如免疫表型、单细胞多组学、适应性免疫受体库)和分子层(如批量转录组、蛋白质组、表观基因组),并讨论跨层映射。
- 归纳建模方法
概述从经典统计模型(线性/正则化回归、生存模型)到机器学习方法(树集成、深度学习)的应用。
- 标准化工作流程
强调从任务定义与队列设计、预处理与特征工程,到验证、重新校准与可重复发布的完整流程的重要性。
- 建立评估框架
- 应用框架分类
- 识别挑战与展望
基于该框架,系统分析当前局限,并提出未来的优先发展方向。
研究亮点:
- 方法学整合
超越单纯罗列生物标志物或模型,提供了一个从队列设计、任务定义、数据预处理、特征工程、模型开发到验证与重新校准的完整方法学框架。
- 强调评估与临床转化
特别强调了模型评估不应仅限于区分度,还必须包括校准和临床净收益分析,并指出了外部验证和跨队列可移植性是当前临床转化的关键瓶颈。
- 明确挑战分类
系统性地总结了阻碍领域发展的三大类挑战:方法学上的批次效应与组成混淆、生物学上的异质性与复杂性、以及证据与报告标准的不完善。
研究结果:
- 日历年龄时钟
可进一步分为基于免疫表型和PBMC转录组的系统水平时钟,以及基于血浆蛋白质组和IgG糖基化的体液分子时钟。它们预测实际年龄,其输出结果“Δ免疫年龄”被解释为相对于同龄人的加速或减速。
- 风险时钟
直接以临床终点(如死亡率、衰弱)进行训练,其输出应解释为风险评分而非“年龄差”。例如,iAge 是一种炎症网络风险时钟,其评分与衰弱、血管老化等临床表型相关。
- 细胞/谱系时钟
关注“时间”在免疫系统内的编码位置,例如特定T细胞亚群的表观遗传状态或TCR/BCR受体库的多样性模式。这类时钟有助于将系统水平读数分解为可解释的细胞组分。
- 当前代表性时钟
(如IMM-AGE, iAge)在输入模态、建模方法和预测目标上各不相同,捕获了免疫衰老的不同维度。
研究总结:
- 主要挑战
- 方法学
批次效应、平台差异、细胞组成混淆是主要干扰;模型可能学习技术噪声而非生物学信号;外部验证与重新校准不足限制跨队列可移植性。
- 生物学
免疫衰老具有高度异质性,受遗传、性别、CMV感染等影响;克隆造血等过程会混淆年龄信号;炎症与代谢网络紧密耦合,难以分离纯粹的免疫衰老信号。
- 证据与标准
多数证据为回顾性关联研究,缺乏前瞻性干预验证;报告质量参差不齐,存在数据泄漏风险;对模型校准和临床效用的评估常不充分;“免疫年龄”的解读与传播存在误区。
- 未来方向
- 多模态整合与深度模型
结合多组学与临床数字数据,构建更具泛化能力和解释性的模型,探索免疫数字孪生框架。
- 组织与细胞类型特异性时钟
超越外周血,开发针对特定免疫细胞谱系和组织微环境的时钟,以定位衰老信号的起源。
- 干预导向的时钟与因果路径
将时钟与可测试的机制通路(如炎症、免疫代谢)和干预研究(如疫苗接种、生活方式)相结合,推动从相关性指标向 actionable 生物标志物转变。
- 建立共享的“免疫年龄”语言
需要通过方法学严谨性、标准化报告、跨队列重新校准机制以及审慎的解读沟通,推动建立可验证、可比较的免疫年龄度量体系。
结果译文:
当前的免疫衰老时钟异质性很强,在输入数据类型和建模选择上差异显著 [14,23,73,97]。输入范围从高维免疫表型到PBMC转录组、外周蛋白组、适应性受体库以及单细胞表观遗传特征 [71,77,89,105]。仅根据模态(例如,PBMC、蛋白质组学或T细胞聚焦的读数)对时钟进行分组,掩盖了它们是解决类似问题还是针对不同目标 [14,21]。一种更实用的方法是根据预测任务对时钟进行分类——即,按时间顺序排列的年龄、临床风险/结局,或编码细胞状态或克隆结构中时间的细胞/谱系水平时钟 [19,68,82]。全血转录组年龄可作为系统水平的参考,将免疫衰老时钟定位在任务-模态-模型框架内。每个时钟最能回答哪个问题,以及它们在不同场景下如何相互补充 [24,71]?关键的代表性时钟及其方法论特征见表2。
1.1 日历年龄驱动的年龄时钟:免疫时间顺序
按时间顺序年龄监督的免疫衰老时钟根据时间顺序年龄尺度进行校准;Δ免疫年龄(预测值减去时间顺序年龄)被解释为与年龄匹配的同伴相比的相对加速或减速 [19,74]。现有研究主要遵循两个轨迹:(i)基于免疫表型和PBMC转录组的系统水平时钟,以及(ii)利用血浆蛋白质组学和IgG糖组学的体液输出导向时钟 [13,14,104]。
在由免疫表型和PBMC转录组驱动的系统水平上,开创性的IMM-AGE研究从随时间收集的高维免疫表型开始 [13]。该方法定义了一个稳定的免疫年龄轨迹,并将其转化为与基因调控活性相关的可重复使用的功能描述符 [13]。随后,在一个外部人群队列(例如,Framingham队列)中评估了其对全因死亡率的独立预后价值 [13]。关键的方法学特征包括明确的任务定义、在结局关联分析中使用ΔIMM-AGE进行生存信息评估、用于可扩展免疫表型的表达特征表示,以及在Cox框架内优先考虑可解释性和外部有效性的正则化线性模型 [13]。
在PBMC bulk转录组水平上,许多免疫相关基因与年龄存在系统性关联 [71,83]。基于模块的方法已经能够建立稳定的免疫转录组年龄时钟(中位绝对误差~3-5年),并提示了中年转录组的拐点 [24,71]。借助更高质量的PBMC单细胞数据,单细胞免疫时钟可以在亚群水平上解析衰老信号 [23,73]。在scRNA-seq年龄梯度中,与年龄相关的信号通常集中在有限的一组亚群上,包括特定的NK状态、CD8+记忆T细胞和CD14+单核细胞 [63,109]。该信号既反映了它们相对丰度的变化,也反映了它们细胞状态的变化 [105]。在感染性疾病和自身免疫性疾病中,免疫年龄加速的报道为从这些时钟推断出的“加速衰老”提供了细胞生物学层面的解释 [23,101]。
相比之下,体液分子时钟利用源自血浆蛋白和IgG糖基化的循环分子网络 [107]。在大型队列(如英国生物银行)上训练的蛋白质组年龄时钟显示出强大的时间顺序年龄预测能力,并在独立队列中具有普适性 [24,77]。虽然基于时间顺序年龄进行训练,但蛋白质组年龄加速已与多个队列中的慢性病负担和全因死亡率相关联,支持其作为风险相关标志物的应用 [14,77]。梯度提升树通常用于对高维蛋白质组特征进行建模 [77,78]。在生存分析中,Δ年龄可以与结局(例如,风险比)相关联,但应明确其定义和校准,并在需要时重新估计,以确保跨队列的可比性 [21,82]。IgG糖组学时钟(例如,GlycanAge)捕获了与年龄相关的Fc N-聚糖谱变化,并产生与代谢和炎症表型相关的Δ年龄测量值,包括生活方式干预后的变化 [104,107]。
总的来说,由日历年龄监督的免疫年龄时钟解决了两个相关问题。一个从外周血免疫组成/状态推断免疫年龄,而另一个则通过循环分子读数反映免疫衰老 [13,73]。前者非常适合对免疫功能及其对感染或疫苗接种反应的机制解释,而后者则支持大规模队列的快速、高维分析 [63,109]。两者都受到外周血采样的限制,可能无法反映组织驻留的免疫衰老 [5]。
1.2. 以死亡率和多病为结局的风险时钟型免疫时钟
风险时钟直接针对临床终点(例如,死亡率、虚弱、多病)进行训练,而不是针对时间顺序年龄 [14,21]。因此,其输出应被解释为终点锚定的风险评分(或预测风险/概率),而不是“年龄差距”。年龄差距的语言仅适用于按时间顺序年龄监督的模型,除非风险评分被明确重新校准到年龄样尺度,并具有明确的假设和外部验证 [32,60]。
iAge 就是一个炎症网络风险时钟的例证。利用来自8-96岁参与者的炎症/免疫蛋白质组学面板,深度模型估计出一个“炎症年龄”,该年龄与多重用药、虚弱和血管老化相关,并可预测心血管事件和死亡率。归因分析突出了CXCL9;后续的实验工作将CXCL9信号传导与内皮功能障碍和动脉僵硬度联系起来,说明了时钟 → 通路 → 机制的工作流程 [14]。相关的蛋白质组学研究通常先训练一个年龄时钟,然后在生存模型中分析年龄加速,或者直接在临床终点上进行监督。对于风险时钟,评估应强调区分度和临床效用(C指数、时间依赖性AUC、决策曲线分析),并谨慎解释Δ年龄:它可能反映的是一个风险连续谱,而非衰老的速度 [93,96,99]。
1.3. 基于细胞状态和适应性受体库的细胞/谱系时钟
与年龄时钟和风险时钟相比,细胞/谱系时钟强调“时间”在免疫系统中是如何编码的——即,哪些谱系/克隆携带与衰老相关的信号,以及这些信号如何在细胞分辨率下以表观遗传状态或受体库的形式存储。这些时钟有助于定位免疫衰老在特定谱系和克隆中的表现 [23,63,73]。
在T细胞谱系中,多组学研究一致报告,随着年龄增长,初始T细胞减少,记忆和终末分化群体相对扩增 [109,110]。在不同组织中,PD-1+TOX+CD8+耗竭样亚群可能很突出,并且通常与慢性局部炎症和代谢稳态受损有关 [63,109]。与这些观察结果一致,基于单细胞PBMC的免疫时钟也突出了特定的年龄相关亚群,包括NKG2C+GZMB+XCL1+ CD8+记忆T细胞,作为跨队列对年龄敏感的稳健亚群 [109]。
TCR/BCR库驱动的“库结构”时钟关注适应性免疫库的多样性和克隆扩增模式 [69,89]。老年人通常表现出多样性降低、优势克隆扩增以及公共克隆型的转变 [35,111]。通过将相似的序列聚类成功能单元,这些单元的多样性和扩增可以映射到年龄或疾病状态,从而实现基于库的年龄或风险尺度。此外,像MiAIRR这样的元数据标准通过支持可互操作的库注释,促进了跨队列的协调和外部验证 [70,105,111];这反过来又支持了将免疫暴露、库结构与下游临床结果联系起来的综合分析。
细胞和谱系时钟之所以有价值,是因为它们能够将聚合的系统水平读数分解为可解释的细胞组分 [105]。这种分解使研究者能够量化加速的免疫衰老,识别贡献最大的谱系,并在适用的情况下推断扩增克隆的分裂历史——从而为谱系靶向干预和组织特异性机制研究提供基础 [63,103,112]。
最近的研究勾勒出一个可行的免疫衰老时钟框架,涵盖了任务定义、模态选择、建模和评估 [32,60,61]。基于血液的标志物可以重复地推导出来,并与发病率和生存率相关联,但将其应用于多样化的现实世界场景仍然是一个关键障碍 [21,77,82]。在方法论上,高维免疫分析放大了批次/平台效应、组成混杂以及非线性模型对特征选择和数据分割的敏感性 [22,41,113]。在生物学上,免疫衰老是异质性的,并受到遗传、性别、慢性炎症和克隆性造血的影响,这使得任何单一的标量评分都变得复杂。最后,不一致的报告和主要来自相关性研究的证据限制了临床可操作性 [102,114]。图3总结了这些障碍。
4.1. 方法论挑战:批次效应、组成混杂和跨队列可迁移性
首先,批次效应和跨平台变异性是不可避免的。免疫衰老时钟主要基于来自不同队列和平台的多组学数据。流式细胞术和质谱流式细胞术数据通常使用不同的抗体panel在不同实验室生成,而转录组学和蛋白质组学分析可能依赖于异质的测序平台或基于质谱的工作流程 [67,115,116]。采样时间、分析前处理、处理方案和冷冻保存条件的差异都可能引入系统性偏差 [85,116]。批次校正方法(如ComBat和Harmony及相关变体)可以在一定程度上减弱明显的批次驱动聚类 [46,84,117]。然而,其性能仍然取决于指定的协变量和有效样本量。遗漏关键协变量,或将真实的年龄相关生物学误判为技术批次效应,都可能留下残留的虚假差异或过度校正而削弱真实信号 [85,116]。这种不确定性在模型开发过程中可能被掩盖,但在外部应用时会表现为可迁移性差 [30,33,50]。
细胞组成混杂通常与批次效应同时发生。全血和PBMC bulk组学时钟本质上是混合模型,同时捕获了细胞内在的分子漂移和亚群比例的变化 [90,118]。反卷积可以提供帮助,但对于稀有亚群、急性炎症扰动或治疗诱导的重塑,其性能会下降 [91,108]。因此,在感染或癌症治疗期间,时钟输出可能反映的是短暂的免疫激活,而非潜在的衰老。虽然非线性模型可以捕获复杂的结构,但它们对数据分割和特征选择很敏感 [55,61]。许多研究报告了良好的内部误差或C指数,但缺乏独立的多中心验证;分层外部验证和重新校准仍然不常见 [29,30]。
最后,即使超越了统计性能,跨时钟的可比性仍然有限。像IMM-AGE、iAge、蛋白质组年龄时钟和基于IgG糖组学的时钟可以应用于同一个体,但捕获的是不同的免疫维度 [13-15,77,107]。因此,“加速”的方向和幅度可能在不同时钟间存在差异 [81]。缺乏一个公认的框架来比较这些时钟在特定任务上的“互补性”和“冗余性”,并控制模型复杂性和输入模态变化,这也阻碍了它们在现实应用中的并行解释 [55,96,119]。为了便于跨时钟的并行解释,表3总结了跨队列异质性的主要来源和推荐的缓解工具。
4.2. 生物学挑战:异质性、CHIP 和炎症-代谢网络
方法论上的挑战主要涉及测量和分析选择,而生物学挑战则反映了潜在免疫系统的内在复杂性。免疫系统衰老受到遗传背景、性别、激素、长期感染(如CMV)、生活方式和环境的影响 [15,123,124]。同龄人之间的免疫谱差异可能与年龄相差十岁或更大的人之间的差异一样大 [23,110]。主要来自单时间点采样的时钟通常无法区分终生升高的炎症基线、近期感染暴露和短期应激反应对观察到的读数的贡献。没有纵向轨迹和机制锚点,“加速免疫衰老”可能被解释为一个复合指标,而不是一个特定的可调节通路 [106,125]。
潜能未定的克隆性造血(CHIP)提供了一个显著的例子。随着年龄增长,携带反复出现的驱动突变(例如,DNMT3A和TET2)的造血克隆可能发生优先扩增,逐渐改变血细胞组成,并导致慢性、低度炎症 [27,28]。CHIP既增加了患血液系统恶性肿瘤的风险,也增加了患动脉粥样硬化和心脏问题的风险。在多个老年人队列中,这种信号可以作为一个潜在层出现,无法从单一测量中直接观察到,但通过整合建模变得明显 [27,28]。如果在时钟构建分析过程中未发现或区分CHIP,那么模型学习到的年龄信号可能对应于拟合CHIP状态,而不是更普遍的免疫衰老。这为后续计划在老年人群中广泛使用的时钟带来了系统性偏倚的来源 [27,28]。
炎症、内分泌调节和代谢在衰老过程中紧密耦合,使得免疫年龄信号与心脏代谢功能障碍、肌肉减少症和认知衰退的分离变得复杂 [6,72]。以炎症和蛋白质组信号为主的时钟通常显示出与多重用药和死亡风险的强关联,但可能部分捕获了系统性虚弱和免疫代谢失调,而不仅仅是免疫特异性功能障碍。然而,目前尚不清楚Δ年龄中有多少反映了免疫特异性功能障碍,又有多少反映了更广泛的系统性虚弱和代谢失调。因此,升高的评分在疾病和治疗环境中的临床意义可能取决于具体情况,这强调了任务特异性验证和校准的必要性 [14,28]。
证据水平与报告标准之间的不对称性是阻碍免疫衰老时钟临床转化和更广泛接受的另一个障碍。迄今为止,大多数证据来自回顾性分析或对最初为其他目的生成的数据集进行的二次建模。这些研究将免疫年龄差距与死亡率、虚弱或多病等结局联系起来,但结论通常是特定于队列和终点的。虽然对产生假设有参考价值,但这种关联并不能自动产生一个可操作的临床指标。临床或公共卫生的采纳需要多中心前瞻性研究,这些研究具有预设的时钟终点和决策阈值——这种证据仍然稀缺 [59,126,127]。
报告质量也参差不齐。数据分割并不总是独立的,特征选择和超参数调整可能不局限于训练数据(存在信息泄露风险),而“外部验证”有时是在仅存在微小分布偏移的队列上进行的;这些细节常常被低估 [33,57]。模型校准状态、阈值选择依据、决策曲线分析以及潜在的临床净收益评估常常缺失或仅在附录中简要讨论。没有标准化的报告,免疫衰老时钟难以重现,也难以纳入指南或共识声明 [55,56]。
解释和沟通也带来了额外的风险。诸如“免疫年龄比同龄人老5岁”这样的说法,作为模型和人群特异的相对位置可能在技术上是有效的,但非专业人士可能会将其解释为确定的生物学加速。在没有干预证据的情况下传递此类读数可能会增加焦虑并鼓励不必要的检测或治疗升级。另一方面,如果有干预证据,我们需要注意不要仅凭一个时钟读数就作为决策的唯一依据。这些问题很少被明确讨论,但对于负责任的现实部署至关重要 [59,127]。
总之,从方法、生物学和证据的角度来看,当前的免疫衰老时钟仍处于“快速增长但未趋同”的状态。解决这些局限性——并克制对时钟输出的解读——将是在超越队列和模型特定最优值的必要步骤。这将使我们更接近于拥有一个真实的、可比较的、可重复的、可理解的数字系统来测量免疫系统的年龄。
更多结果和补充图表:doi: 10.3390/cells15050421
长按二维码关注我们,用最短的时间和最高的效率学习更多数据分析方法!
扫描上方二维码或登录平台官网后添加CNSknowall客服微信咨询!官网地址:
https://cnsknowall.com
CNSknowall:24年最新问世的遥遥领先的科研数据(0代码生信+统计学)分析平台,同时含有机制图模块+汉化版Pubmed融合Deepseek高效筛选目标文献+SCI文献例句/语料检索模块+OPenAI官方GPT接口,>500款CNS级别图表皆可一秒内一键出图,登录即秒变数据分析大神,体验前所未有的便捷数据分析之旅,开启科研天骄之路!
可向下滑动批阅!