传统疼痛研究往往局限于单个脑区或分子,难以描绘神经病理性疼痛在完整神经轴上的全景变化。而本文通过整合高分辨率蛋白质组学与多区域比较分析,在坐骨神经、脊髓和眶额皮层中系统解析了神经免疫与多巴胺能信号的协同改变。这种“多节点组学+功能验证”双保险策略,不仅锁定S100蛋白等关键靶点,更揭示了情感-认知环路对疼痛的调控机制,为你提供更全面、更易转化的疼痛机制研究新范式。
今天给大家解读的一篇文章,是2月发表在《Cells》上的题目为“Integrative Proteomics Reveal Neuroimmune and Dopaminergic Alterations Across the Nociceptive Neuraxis in Neuropathic Pain.”这篇文章旨在阐明神经病理性疼痛(NP)在整个痛觉传导神经通路(从外周神经到中枢皮层)中的分子改变。通过建立CCI大鼠模型并确认其出现痛觉超敏、异常性疼痛及运动功能障碍后,研究人员采集了损伤侧的坐骨神经(SN)、脊髓(SC)和眶额叶皮层(OFC)组织进行高通量蛋白质组学分析。通过生物信息学方法鉴定差异表达蛋白(DEPs)并分析其功能通路,再使用免疫荧光和ELISA对关键发现进行验证。文章最终提出,NP由神经免疫激活和涉及多巴胺信号传导的神经调节适应共同驱动,影响疼痛的感觉和情感成分。(请持续关注我们,每天为您解读最新见刊的文献!)想薅生信资料羊毛?直接在对话框回复 “资料”,免费领取干货大礼包!
不想做实验,没数据,还想要快速发表文章,没问题的!公共数据库就是我们的数据宝藏!没思路不用担心,作为专业的生信团队,我们很乐意为你们效劳,提供研究路线设计和数据挖掘分析,扫码联系我们吧!


团队成员合影(位于上海陆家嘴中心,可随时预约参观)
题目:《整合蛋白质组学揭示神经病理性疼痛中痛觉传导神经通路的神经免疫和多巴胺能改变》Integrative Proteomics Reveal Neuroimmune and Dopaminergic Alterations Across the Nociceptive Neuraxis in Neuropathic Pain
发表期刊:Cells
影响因子:5.2
研究背景:神经病理性疼痛(NP)影响全球约7-10%的人口,约35%的患者对经典镇痛药无效。其分子机制尚未完全阐明。传统研究多聚焦于感觉通路和脊髓水平的下行抑制通路,而对疼痛的情感维度及整合感觉与情感的高级皮层区域关注不足。眶额叶皮层(OFC)是整合疼痛情感和认知成分的关键枢纽,并与奖赏环路相连。本研究提出“痛觉传导神经通路”的概念,旨在通过涵盖外周(SN)、脊髓(SC)和高级皮层(OFC)的蛋白质组学分析,系统揭示NP的分子基础,以发现新的治疗靶点。
CNSknowall 平台 Pubmed+AI 快速提炼全文要点
研究思路:
- 模型建立与验证
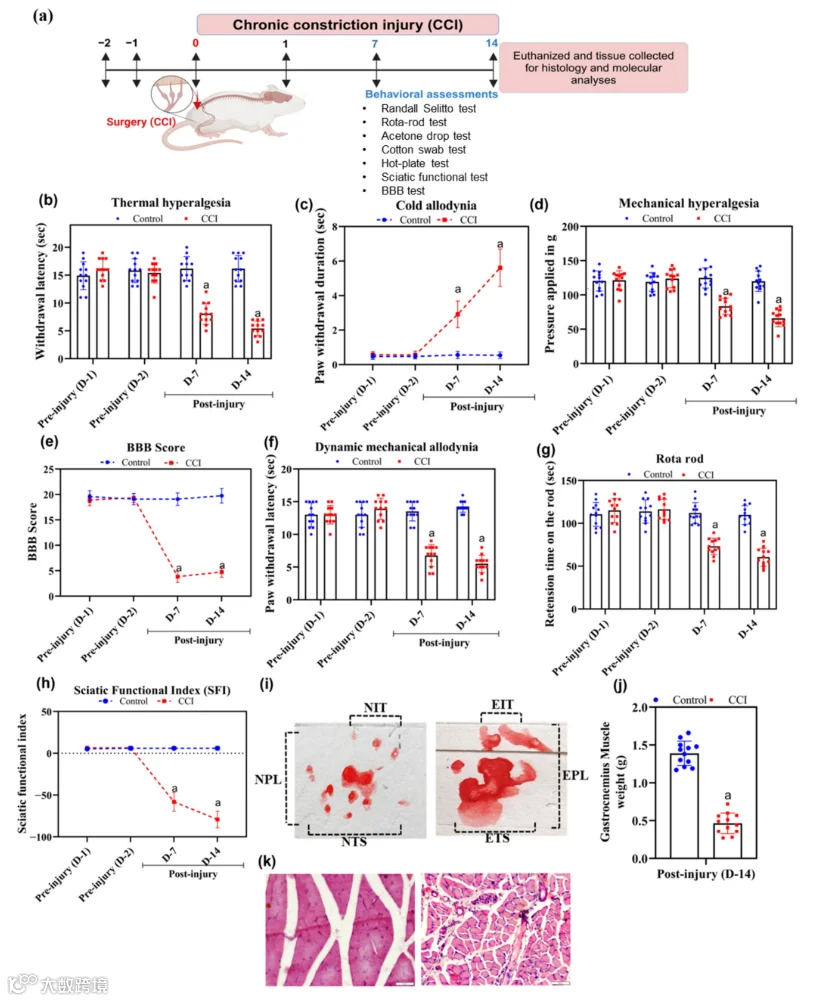
采用雄性Wistar大鼠建立CCI模型,通过一系列行为学测试(热板、丙酮、Randall Selitto、棉签、Rota rod、SFI、BBB)在术后第7和14天评估痛觉超敏、异常性疼痛及运动功能损伤,并通过腓肠肌重量和组织学确认肌肉萎缩。
- 样本采集与蛋白质组学分析
于术后第14天(慢性NP稳定期)取同侧SN、SC和OFC组织。使用LC-HRMS技术进行非标记定量(LFQ)蛋白质组学分析。
- 数据分析
1)鉴定各组织中CCI组与对照组的差异表达蛋白(DEPs)。
2)对DEPs进行基因本体论(GO)和京都基因与基因组百科全书(KEGG)通路富集分析。
3)使用Ingenuity Pathway Analysis(IPA)进行经典通路、上游调节因子和分子网络分析。
4)对CCI组特有的蛋白进行单独的功能和通路分析。
5)比较SN、SC和OFC三者之间共同的经典通路。
- 实验验证
通过免疫荧光检测胶质细胞标志物(GFAP, Iba-1)和S100蛋白(S100B, S100A8)的表达与定位;通过ELISA检测组织中的促炎细胞因子(TNF-α, IL-6)和多巴胺(DA)水平。
研究亮点:
- 全局视角
首次在CCI模型中,对涵盖外周神经、脊髓和关键前脑皮层(OFC)的完整痛觉通路进行同步蛋白质组学分析。
- 发现共性靶点
S100蛋白家族(S100A8和S100B)在SN、SC和OFC中均显著上调,并与胶质细胞活化(GFAP, Iba-1)及促炎细胞因子(TNF-α, IL-6)水平升高同步,提示其作为NP的跨区域神经免疫枢纽分子。
- 揭示情感环路机制
在负责情感和认知处理的OFC中,发现多巴胺调节蛋白上调和多巴胺能神经传递的正向调控,首次在蛋白质组水平将多巴胺能奖赏相关镇痛环路与NP的慢性化联系起来。
- 数据整合深入
结合差异表达蛋白(DEPs)筛选、GO/KEGG功能富集、IPA通路分析以及免疫荧光、ELISA验证,构建了一个从分子到表型的整合性分子图谱。
研究结果:
- 行为与病理表型
CCI大鼠表现出显著的热痛觉超敏、冷异常性疼痛、机械痛觉超敏和动态机械异常性疼痛,同时伴有运动协调能力下降、坐骨神经功能指数(SFI)降低以及腓肠肌萎缩和炎性浸润。
- 蛋白质组学概览
1)SN
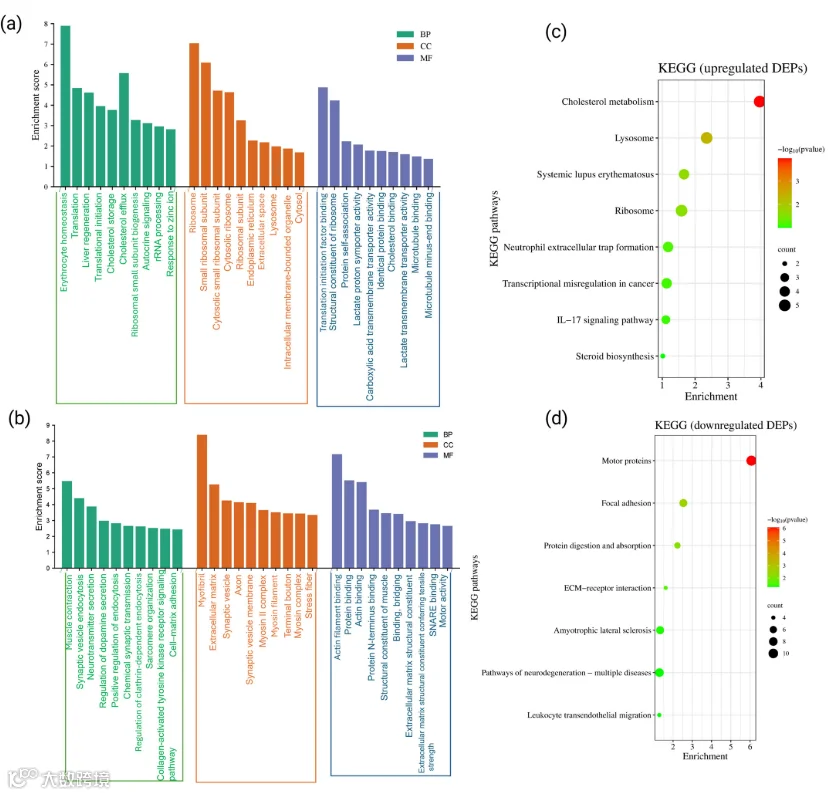
鉴定到3875个DEPs。上调蛋白富集于红细胞稳态、翻译、胆固醇代谢、核糖体生物合成等;下调蛋白与肌肉收缩、突触囊泡内吞、神经递质分泌等相关。KEGG提示炎症(如IL-17)、胆固醇代谢和核糖体通路上调;黏着斑和ECM-受体相互作用通路下调。
2)SC
鉴定到4428个DEPs。上调蛋白涉及突触可塑性(如长时程增强)、突触传递、钙离子结合等;下调蛋白与线粒体呼吸链复合物I组装、氧化磷酸化等相关。KEGG显示补体和凝血级联、逆行内源性大麻素信号等通路上调;阿尔茨海默病、帕金森病等神经退行性疾病通路下调。
3)OFC
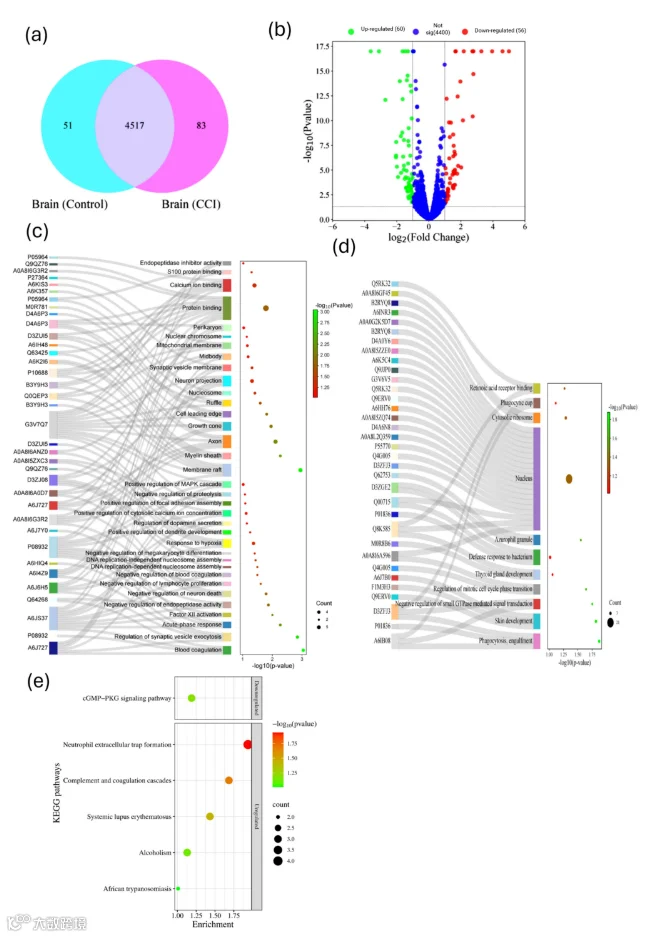
鉴定到4517个DEPs。上调蛋白与调节突触囊泡胞吐、急性期反应、负调控神经元死亡、调节多巴胺分泌等有关;下调蛋白涉及小GTP酶信号负调控等。KEGG显示中性粒细胞胞外陷阱形成、补体和凝血级联等通路上调;cGMP-PKG信号通路下调。
- 跨区域共同通路(IPA分析)
SN、SC和OFC共享的典型通路包括中性粒细胞胞外陷阱形成、SNARE信号、S100家族信号、NMDA受体信号、血清素受体信号等,强调了神经免疫和突触可塑性改变的系统性。
- 关键验证结果
1)神经免疫激活
免疫荧光证实CCI后SN、SC和OFC中星形胶质细胞(GFAP+)和小胶质细胞(Iba-1+)显著活化,形态向反应态转变。
2)S100蛋白普遍上调
S100A8和S100B在SN、SC和OFC中的蛋白表达均显著增加。
3)炎症因子升高
ELISA显示CCI大鼠SN、SC和OFC中促炎细胞因子TNF-α和IL-6水平升高。
4)多巴胺能改变
ELISA显示CCI大鼠SC和OFC中的多巴胺(DA)水平显著升高。蛋白质组数据也显示OFC中多巴胺分泌调节和多巴胺能神经传递正调控相关蛋白富集。
研究总结:
-
NP涉及从外周神经到前脑皮层的系统性分子重编程,而非孤立的局部事件。
-
神经免疫激活是贯穿该通路的中心机制,表现为胶质细胞活化、S100蛋白(S100A8/S100B)上调和促炎细胞因子释放,共同营造了慢性的炎症微环境。
-
多巴胺能系统在高级皮层(OFC)被显著调动DA水平升高及相关蛋白上调,表明奖赏和动机相关环路参与了NP的适应过程,这可能与疼痛的情感/认知维度处理相关。
-
研究提出了一个由神经免疫激活和神经调节适应(尤其是多巴胺能信号)共同驱动的痛觉传导神经通路模型,该模型交互作用,共同影响疼痛的感觉和情感成分。
-
潜在治疗靶点研究结果提示,靶向胶质细胞衍生的S100蛋白(如S100A8, S100B)及其下游信号(如RAGE受体),以及调节多巴胺能系统,可能是未来综合管理NP(同时针对感觉和情感症状)的新策略。
结果译文:
1.CCI对大鼠感觉和运动功能的影响
行为学测试在CCI手术后的第7天和第14天进行,以确认大鼠神经病理性疼痛(NP)的发展(图1a)。已有报道指出,CCI动物模型可引发对各种刺激的痛觉过敏和异常性疼痛,并伴随小腿肌肉萎缩[50]。同样,在我们的研究中,通过双因素方差分析(two-way ANOVA)及事后检验的统计分析显示,从第7天起与对照组相比,实验组大鼠出现了热痛觉过敏、冷异常性疼痛、机械痛觉过敏以及动态机械异常性疼痛(p < 0.05)(如图1b–f所示)。周围神经损伤导致大鼠在热板上放置时撤退潜伏期显著缩短(图1b),同时在受伤足底滴加丙酮后足部撤退持续时间增加(图1c);上述变化在组别方面分别为[F(1,88) = 117.4; p < 0.05] 和 [F(1,88) = 351.4; p < 0.05],时间方面分别为[F(3,88) = 33.98; p < 0.05] 和 [F(3,88) = 114.3; p < 0.05],以及它们的交互作用分别为[F(3,88) = 48.21; p < 0.05] 和 [F(3,88) = 135.8; p < 0.05]。此外,还观察到在Randall-Selitto试验中,大鼠承受施加于足背的压力阈值显著降低(图1d),且该组的BBB评分也显著低于对照组([F(1,88) = 1084; p < 0.05],时间[F(3,88) = 336.4; p < 0.05]及其交互作用[F(3,88) = 342.7; p < 0.05],如图1e所示)。CCI术后坐骨神经(SN)损伤后,大鼠出现了动态机械异常性疼痛(图1f),并且还表现出运动不协调及肌力下降,表现为在旋转棒上停留时间减少(图1g),组别差异为[F(1,88) = 66.93; p < 0.05],时间差异为[F(3,88) = 35.52; p < 0.05],以及其交互作用[F(3,88) = 31.31; p < 0.05]。此外,实验大鼠在第7天和第14天的SFI评分分别降至−58.08 ± 11.34和−79.25 ± 9.87,而术前为6.58 ± 1.31,提示运动不协调的发生(图1h,i)。另外,CCI损伤后,CCI组同侧腓肠肌重量显著减轻(即0.46 ± 0.136 g),相较于对照组(即1.39 ± 0.161 g),表明存在肌肉萎缩(图1j)。类似地,对照组腓肠肌组织学分析显示形态均一,骨骼肌纤维呈密集排列的多边形形状,并伴有周边椭圆形细胞核。相反,CCI组则显示出肌浆碎裂、单核细胞浸润,以及内膜间隙增宽的现象(图1k)。
2.坐骨神经中差异表达蛋白(DEPs)的定量及上调和下调蛋白的鉴定
为了阐明周围神经损伤后脊髓(SN)中的分子变化,我们对对照组和CCI诱导的NPrats进行了基于高分辨率质谱的蛋白质组学分析。蛋白质组谱分析在对照组和CCI组的SN中分别鉴定出3994种和4388种蛋白质。进一步地,在比较CCI组与对照组时,共鉴定出3875种差异表达蛋白(DEPs),占总蛋白数的86%,如图S1a所示的维恩图所示。我们还观察到,有513种(11.4%)蛋白质仅在CCI大鼠中表达,而119种(2.6%)蛋白质仅在对照组中表达。此外,在这些DEPs中,显著上调和下调的蛋白质在火山图中被识别并展示(图S1b)。在CCI后第14天,SN中共有91种蛋白质显著上调,而168种蛋白质显著下调(图S2)。CCI后SN中上调最多的前10种蛋白质分别为:S100钙结合蛋白A8(Calgranulin A;A6J6Q3)、RIKEN cDNA样蛋白(A6JIC0)、锌指蛋白551(A0A8I5Y616)、转位子相关蛋白亚基γ(F1M7T6)、防御素NP-4(A6IW99)、NPC细胞内胆固醇转运蛋白1(A0A8I6GB30)、蛋白磷酸酶1G(F1LNI5)、核糖体蛋白S6激酶(A6HZI4)、组蛋白H3(D3ZJ08)和组织蛋白酶D(Q6P6T6)(见表1)。同样,在这168种统计学上显著下调的蛋白质中,前10种蛋白质分别为:非特异性丝氨酸/苏氨酸蛋白激酶(A0A0G2JUP3)、LON肽酶N端结构域和RING指蛋白1(A0A0G2K313)、白蛋白、异构体CRA_b(A6KKG1)、髓鞘碱性蛋白(MBP;A6K5J1)、糜蛋白酶1(A6KH67)、核孔膜糖蛋白210(A6IB90)、锌指蛋白831(A0A0G2JZX4)、角蛋白83(A7M746)、半胱氨酸和甘氨酸丰富蛋白1(A6ICG2)以及RCG61079异构体CRA_a(A6JKY8)(表1)。
3.坐骨神经在CCI损伤后上调和下调表达蛋白的功能注释
为了全面评估CCI后SN中DEPs的功能影响,我们使用DAVID生物信息学资源对上调和下调蛋白进行了GO分析,结果如图2a、b所示。在上调的DEPs中,GO注释覆盖率较高,其中94.3%映射到生物过程(BP),93.1%映射到细胞组分(CC),89.7%映射到分子功能(MF)。在显著富集的BP方面,SN中上调的蛋白主要与红细胞稳态、蛋白质翻译、肝脏再生、胆固醇储存与外排、核糖体小亚基生物发生、自分泌信号传导、rRNA加工以及对锌离子的反应相关(图2a)。这些过程表明受损SN微环境中代谢活性增强、核糖体生物发生增加以及稳态补偿机制的激活。在细胞组分层面,上调蛋白主要与细胞内细胞器如核糖体、内质网(ER)、溶酶体及细胞内膜包被细胞器相关。此外,在分子功能方面,上调蛋白富集于翻译起始因子结合、蛋白质自我关联、乳酸:质子同向转运体活性、羧酸跨膜转运体活性、胆固醇结合、乳酸跨膜转运体活性以及微管结合等类别。胆固醇结合蛋白的富集尤为重要,因为胆固醇丰富的脂筏(也称为炎性脂筏)的形成增加已被证明与临床和临床前模型中的神经炎症相关[51,52]。因此,在我们的数据集中,SN中上调的蛋白似乎有助于神经炎症信号传导以及由突触可塑性驱动的疼痛慢性化。
同样,对于下调的DEPs,下调蛋白的GO覆盖度也相当可观:生物过程(BP)为89.0%,细胞组分(CC)为96.6%,分子功能(MF)为82.2%。在显著富集的BP术语中,肌肉收缩、突触囊泡内吞、神经递质分泌、多巴胺分泌调节、内吞作用正调控、化学突触传递、网格蛋白介导的内吞、肌节组织、胶原激活的酪氨酸激酶受体信号传导以及细胞-基质黏附在SN中显著富集(图2b)。此外,MF分析显示,下调蛋白在以下功能注释中富集:蛋白质结合与桥接、肌动蛋白丝结合、肌肉结构组分、赋予张力强度的细胞外基质成分以及运动活性(所有GO术语的p值均<0.05)。除此之外,根据GO分析,参与SNARE结合的蛋白也呈下调趋势。为进一步确定CCI后SN中统计学上显著上调和下调的DEPs的潜在生理通路,还进行了KEGG通路分析,并分别展示在图2c和图2d中。根据我们的分析,55.2%的上调DEPs和45.9%的下调DEPs被注释到已知的KEGG通路。在上调蛋白中,最显著富集的KEGG通路包括胆固醇和类固醇生物合成与代谢、炎症信号(例如IL-17信号)、中性粒细胞浸润、溶酶体活性以及核糖体通路(图2c)。这些通路提示损伤神经中免疫反应增强和蛋白质周转增加。相反,下调DEPs的KEGG分析表明,在焦点黏附和细胞外基质(ECM)-受体相互作用等通路中显著富集(图2d)。这些蛋白与结构完整性、细胞-基质相互作用以及细胞骨架支持系统相关。此外,若干下调蛋白还与神经退行性疾病通路和白细胞迁移相关。
4.SN蛋白质组的Ingenuity Pathway Analysis (IPA)
为了深入理解驱动CCI模型中神经病理性疼痛进展的规范通路、疾病关联及上游调节因子,使用SN中的DEP数据集对Ingenuity Pathway Analysis (IPA)进行了分析。基于z评分的前五条显著富集的规范通路包括细胞外基质(ECM)组织、SNARE信号传导、胶原纤维组装、中性粒细胞脱颗粒以及肝星状细胞激活(图S3)。免疫相关信号通路如MHC II类抗原呈递和白细胞介素信号传导也显著富集(p < 0.05)。此外,钙信号传导、神经递质释放、神经细胞黏附分子(NCAM)信号传导和轴突生长也明显观察到。除此之外,胶原生物合成、脂蛋白重塑以及肝脏X受体/视黄醇X受体(LXR/RXR)激活相关的通路在受损的SN中也显著存在,表明广泛的生理重塑。IPA的疾病与功能模块揭示了与神经系统疾病、机体损伤、心血管及心理功能相关通路的显著富集。共识别出16个功能分子网络,其中顶级网络涉及与代谢性疾病、神经系统发育及神经功能相关的基因和蛋白质(表S1)。值得注意的是,离子型NMDA受体(GRIN3A、GRIN1)、PRKAR2A、PRKCE和NDUFS4是该网络中的关键分子(图S4)。
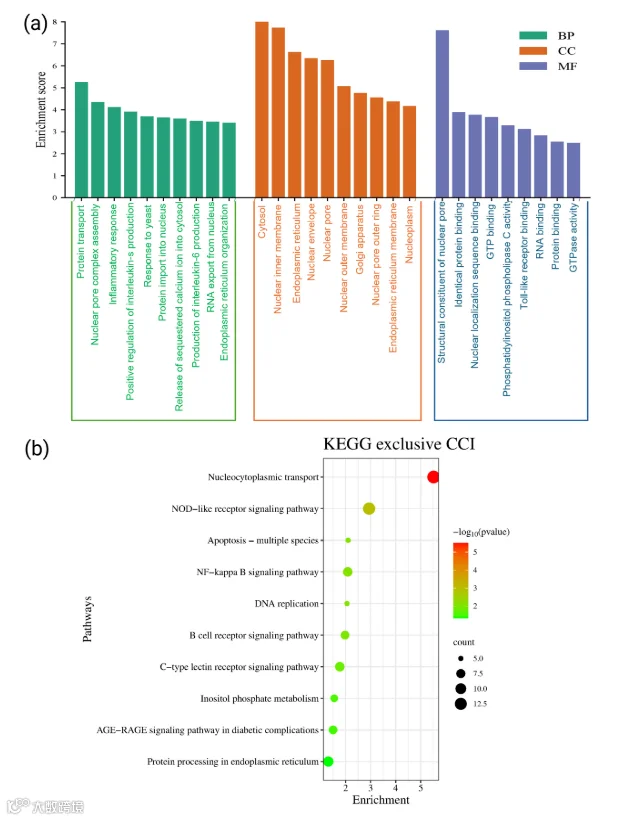
5.CCI组小鼠坐骨神经中特有表达蛋白的基因本体论和通路分析
CCI诱导的神经病理性疼痛大鼠的SN蛋白组学分析揭示了4388种蛋白质的表达,其中513种仅在CCI组中表达。这些独特蛋白质被用于GO和KEGG通路富集分析,以揭示其生物学相关性(图3)。生物过程(BP)的GO分析显示,与炎症细胞因子产生、肿瘤坏死因子(TNF-α)信号传导以及NF-κB转录因子活性的正向调节相关的通路显著富集(图3a)。这些蛋白质还与核质转运机制相关,包括核孔复合体(NPC)组装。其他生物过程包括胞质钙释放和磷脂酰肌醇介导的信号传导。在细胞组分(CC)领域,这些独特蛋白质主要定位于胞质区室、内质网(ER)、核膜、Cajal体及内质网到高尔基体囊泡(图3a)。对于分子功能(MF),富集于核定位序列结合、RNA结合、Toll样受体结合和磷脂酶C活性,表明它们参与了凋亡和免疫激活(所有术语p < 0.05)。同样,KEGG通路分析进一步显示,这些蛋白质参与NOD样受体信号传导、NF-κB和B细胞受体信号传导、凋亡以及AGE-RAGE和C型凝集素受体信号等炎症通路(图3b)。总体而言,这些发现强调了CCI损伤后SN中独特的蛋白质特征涵盖了炎症、细胞信号传导、转录调控和神经元兴奋性的关键调节因子,从而对神经病理性疼痛的病理生理学和适应性反应均有贡献。
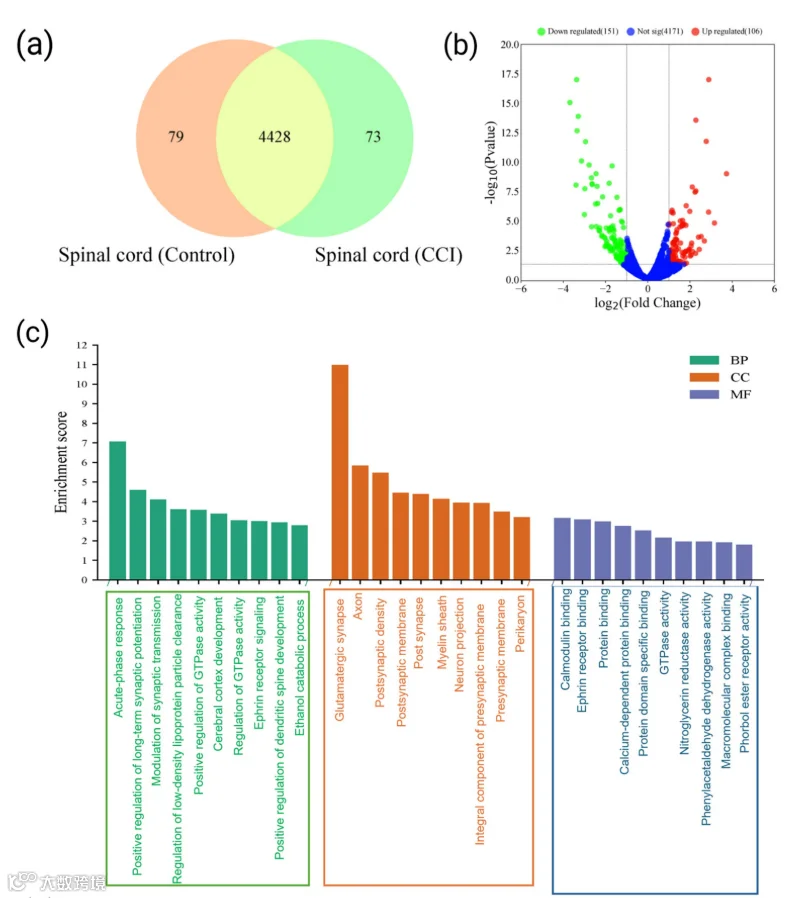
6.差异表达蛋白(DEPs)的定量及脊髓中上调和下调蛋白的鉴定
CCI诱导的神经病理性疼痛(NP)大鼠脊髓(SC)的蛋白质组学分析揭示了蛋白质表达的显著改变。在对照组SC样本中检测到4507种蛋白质,CCI组中检测到4501种蛋白质。其中,4428种蛋白质(96.7%)差异表达(图4a)。在这部分蛋白质中,73种仅存在于CCI组,79种仅存在于对照组。火山图分析(图4b)识别出在CCI术后14天时,SC中106种蛋白质显著上调,151种蛋白质显著下调。这些发现经热图分析(图S5)进一步确认,显示这些蛋白质在CCI组中丰度更高。上调蛋白质中的显著候选蛋白包括胃泌素释放肽、T-激肽原2、突触结合蛋白样蛋白4、S100A8、sequestosome-1和syntaxin结合蛋白1(表2)。这些蛋白质与痛觉感知、炎症、钙信号传导及突触传递相关,可直接或通过释放多种炎症介质促进中枢敏化。相反,最显著下调的蛋白质包括肌浆/内质网钙ATP酶-1、膜相关鸟苷酸激酶(MAGUK)、水通道蛋白-1和SH3结构域结合蛋白1。上述蛋白质分别与钙稳态、脊液调节、细胞骨架动力学及突触稳定性相关。综上,CCI大鼠SC的蛋白质组景观凸显了CCI后SC中独特的蛋白质组改变,并提示涉及促炎信号、钙紊乱及突触重塑的复杂调控网络,共同参与中枢敏化及NP病理生理过程。
7.CCI损伤后脊髓中表达的上调和下调蛋白的功能注释
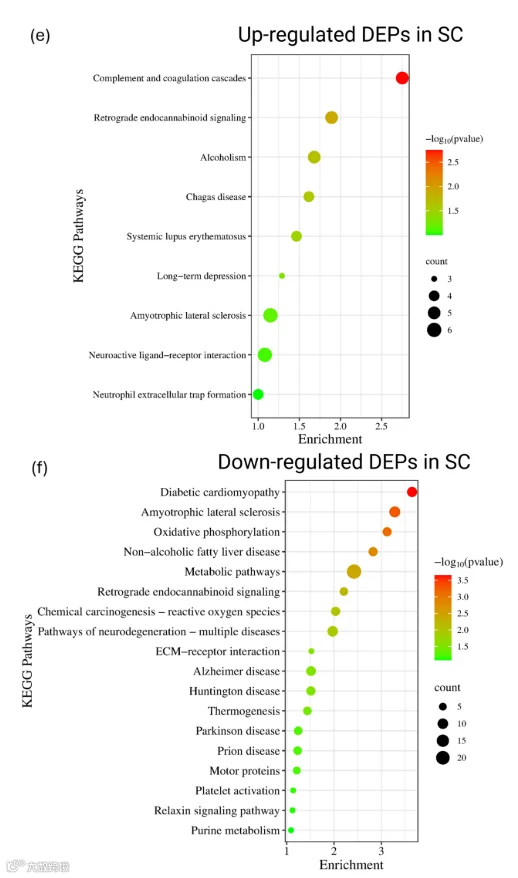
对SC中上调和下调的DEPs进行了GO富集分析,以评估CCI诱导的功能重塑(图4c,d)。在上调的DEPs中,94.2%被注释到BP,95.1%被注释到CC,94.2%被注释到MF。同样,下调的DEPs也表现出广泛的GO覆盖:BP为90.3%,CC为88.8%,MF为76.9%,表明周围神经损伤在SC中引发了广泛的蛋白质组变化。在BP类别中,上调蛋白主要富集于正调控长时程突触增强(LTP)、突触传递、树突棘形态发生、胞外分泌过程中囊泡锚定、eph受体信号传导以及氧化应激反应等过程(图4c)。在CC类别中,它们定位在谷氨酸能突触、树突棘、轴突、髓鞘以及突触前/后膜。在MF本体中,它们涉及钙依赖性结合(如钙调蛋白和S100家族蛋白)、Eph受体结合、磷脂酶抑制剂活性以及RAGE结合。相比之下,下调的DEPs显著富集于线粒体呼吸链复合物I组装、整合素介导的信号传导、过氧化氢反应及碳水化合物分解代谢等BP术语(图4d)。此外,这些蛋白定位于线粒体复合物I、核周细胞质、Ⅰ型胶原三聚体、高尔基体、内质网-高尔基体中间区室以及微管等结构。在MF方面,它们富集于离子通道结合、整合素和微管蛋白结合、血小板源性生长因子(PDGF)结合、FAD结合以及微管相互作用。进一步地,对SC中上调和下调的DEPs进行了KEGG通路分析,以探究CCI后富集的生物通路。上调DEPs的KEGG覆盖率为55.3%,下调DEPs为54.5%。在上调蛋白中,关键富集通路包括补体与凝血级联反应、逆行内大麻素信号传导、长时程抑制,以及与恰加斯病、酗酒症和系统性红斑狼疮相关的通路(p < 0.05;图4e)。此外,补体蛋白如补体成分C9、α-2-巨球蛋白和补体因子B也显著升高,提示周围神经损伤后SC微环境中先天免疫显著激活,并伴随局部促炎细胞因子的急剧增加。相反,下调的DEPs主要富集于氧化磷酸化、神经退行性疾病(如阿尔茨海默病、帕金森病、肌萎缩侧索硬化症、亨廷顿病)、朊病毒病和糖尿病心肌病相关的通路(图4f)。这些结果表明,周围神经损伤后SC中线粒体呼吸活性降低,并且更广泛地抑制了神经保护性和结构信号通路。
8.SC蛋白质组的Ingenuity Pathway Analysis (IPA)
CCI后SC蛋白质组的IPA分析揭示了一个高度互联的分子网络,该网络调控周围神经损伤向持续性神经病理性疼痛(NP)的转变,如图S6所示。对上调和下调的差异表达蛋白(DEPs)进行的经典通路映射识别出五个显著通路:GP1b-IX-V激活、NMDA受体介导的突触后信号传导、EGR2/SOX10依赖的施万细胞髓鞘形成、血小板与暴露胶原的黏附以及细胞外基质(ECM)重排(表3)。
排名最高的通路是GP1b-IX-V信号通路,这是一种由损伤后细胞外基质破坏激活的血小板黏附级联反应。该通路促进磷脂酶C的激活、细胞内Ca2+释放以及趋化因子产量增加,从而导致免疫细胞向脊髓(SC)募集。这些浸润的白细胞与胶质细胞相互作用,增强促炎细胞因子的产生,最终使伤害感受神经元敏感化并持续疼痛信号传递。其他上调的通路包括Ephrin B信号通路和p75NTR受体介导的信号通路,两者均与神经免疫调节和神经元可塑性相关。IPA分析揭示,在CCI大鼠中GABA能信号通路受到抑制,而内源性大麻素突触信号通路被激活。值得注意的是,α-肾上腺素信号通路也被确定为关键介质,参与脊髓轴中的异位神经元放电和胶质细胞激活。在这些发现中,NMDA受体介导的突触后信号传导通路显著富集,其中上调的亚单位包括GRIN1和GRIN3A。此外,为了更深入地了解在SC中表达的映射蛋白的功能,我们进一步分析了这些差异表达蛋白(DEPs)在相关疾病和生物功能中的作用。分析结果证实,SC中映射的DEPs与机体损伤以及神经、心血管和胃肠疾病相关。在生理功能方面,这些显著表达的蛋白质被确定与神经系统发育及其他组织修复相关(表S2)。IPA提出的一个最为显著的网络在研究神经系统疾病、机体损伤及异常和心理障碍中起着关键作用。在此网络中,对神经病理学(NP)起关键作用的分子包括BDNF、CAV1/2、CHRNA7、CNKSR2、CSF3、GABRG2、GNG7、GRIN亚单位、KCNH5、MAP2、NECAB2、RNASE4、SLITRK1、SRC、SUMO2/3和TNC(图S7)。
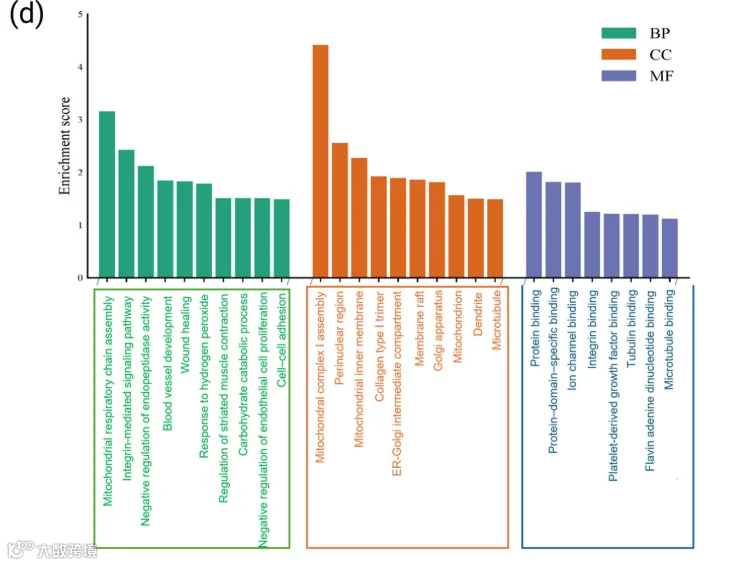
9.CCI组脊髓(SC)中特有表达蛋白的基因本体论和通路分析
SCI术后14天的SC蛋白组学分析鉴定出4501种蛋白质,其中73种仅在CCI组中表达。这十种特有蛋白包括S100钙结合蛋白A9、细胞色素c氧化酶亚单位2、肿瘤坏死因子受体超家族成员21、磷脂酶A2、NADH脱氢酶(泛醌)1α亚复合物、补体C8、蛋白S100-A3、二肽基肽酶9、载脂蛋白C-III和exportin 7。免疫荧光证实S100家族蛋白在CCI大鼠的SN、SC和OFC中表达,提示其在损伤后上调。此外,exportin-7的存在表明microRNA的核质转运发生改变,可能调节损伤应答基因表达。对这73种特有蛋白进行GO富集分析显示,生物过程(BP)涉及甘油三酯代谢、坏死性细胞死亡和蛋白质磷酸化调控。其他富集的BP术语包括肌动蛋白细胞骨架组织、神经发生和线粒体电子传递(图5a)。类似地,细胞组分(CC)分析显示这些蛋白显著定位于胞质溶胶、核膜、线粒体和着丝点。分子功能(MF)术语强调水解酶活性、激酶结合、GTP酶调节活性和孔蛋白活性,反映了它们在膜动态、信号传导和能量代谢中的作用。这些功能体现了NP病理生理过程中膜动态、细胞骨架重塑和脂质信号传导的交集。此外,潜在且统计学显著的KEGG通路将这些特有蛋白与炎症相关脂质信号通路联系起来(图5b)。
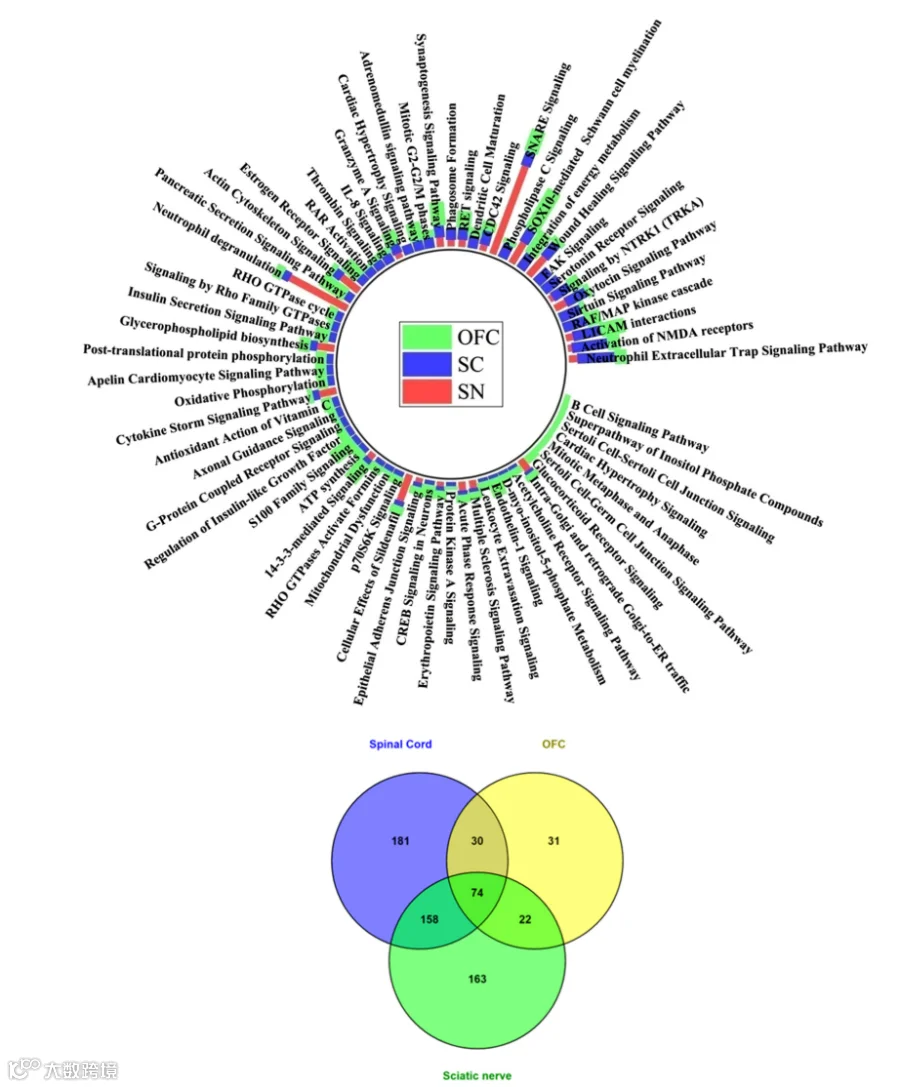
10.外周神经CCI后SN与SC之间的常见规范通路
为了阐明沿所提出的外周-中枢疼痛轴维持NP的共享分子机制,蛋白质组学分析识别出在CCI后背根神经节(SN)和脊髓(SC)中均存在的300条经典通路(图S8a,b)。其中,前十位共享通路包括细胞外基质(ECM)组织、网格蛋白介导的内吞作用、谷氨酸受体信号传导、中性粒细胞胞外陷阱形成、微自噬信号传导、清道夫受体配体结合、血清素受体信号传导、钙信号传导、p75 NTR受体介导的信号传导以及SNARE信号传导(表4)。这些共享通路突显了外周和中枢损伤部位之间广泛的分子重叠。
11.前额叶皮层(OFC)中差异表达蛋白(DEPs)的定量及上调和下调蛋白的鉴定
CCI后对OFC的蛋白质组学分析揭示了显著的大脑皮层蛋白质组重塑,这与在SC和SN中观察到的变化相呼应。对照组和CCI大鼠的OFC中分别鉴定出4568种和4600种蛋白质,其中4517种蛋白质(97.1%)为两组共有(图6a)。值得注意的是,有83种蛋白质(1.8%)仅存在于CCI组,51种蛋白质(1.1%)仅存在于对照组,表明周围神经损伤引发了深刻的蛋白质组变化。火山图分析显示,CCI组中有56种蛋白质显著上调,60种蛋白质下调(图6b),热图聚类分析则显示CCI样本中存在一致的上调趋势(图S9)。在OFC中表达最上调的蛋白质包括RAD21,这是染色质环形成和基因转录所必需的核心凝缩蛋白复合物亚单位;NADPH依赖性3-酮类固醇还原酶(AKR1C18),这是神经类固醇生物合成的关键酶;CD200,一种调节小胶质细胞反应性的免疫调节糖蛋白;碳酸酐酶11(CA11),它催化CO2的可逆水合反应并影响细胞内pH;T-激肽原2和激肽原,是促炎缓激肽的前体;含三核苷酸重复序列18(TNRC18),参与RNA加工;以及非受体型酪氨酸磷酸酶PTPN14,它调节细胞骨架和连接结构的动力学(表5)。相比之下,下调的蛋白质包括黏附型G蛋白偶联受体V1(aGPCRs)、组蛋白H2B类型1、外骨素样糖基转移酶2、NADH-泛醌氧化还原酶链2等。
12.CCI损伤后OFC中表达的上调和下调蛋白的功能注释
为了描绘外周神经损伤对脊上环路的功能影响,我们对大鼠在坐骨神经结扎(CCI)术后第14天眶额皮层(OFC)中表达显著改变的蛋白质进行了全面的GO富集分析(图6)。在上调的差异表达蛋白(DEPs)所覆盖的三个GO类别中,生物过程(BP)、细胞组分(CC)和分子功能(MF)的覆盖率分别为92.5%、92.5%和92.5%。同样,下调的DEPs在BP、CC和MF中的覆盖率分别为89.3%、92.9%和82.1%。就生物过程而言,OFC中上调的蛋白质主要聚集于调控突触功能和细胞存活的过程。最显著的BP术语包括突触囊泡胞吐作用的调节、急性期反应、内肽酶活性的负向调节、神经元死亡的负向调节以及DNA复制依赖的核小体组装。这些过程涉及增强神经递质释放、炎症反应、蛋白酶调控、神经保护及染色质重组。其他富集的过程还包括血液凝固、因子XII激活以及与免疫相关的BP,如巨核细胞分化和淋巴细胞增殖的负向调节,反映了机体试图通过调节CCI后外周免疫细胞浸润或胶质细胞活化来维持稳态。此外,“细胞质钙离子浓度的正向调节”、“多巴胺分泌的调节”和“MAPK级联反应的正向调节”等术语揭示了介导皮层兴奋性和突触可塑性的神经递质传递和细胞内信号传导的变化,从而导致持续性疼痛感觉。同样,在上调蛋白的细胞组分(CC)富集中,发现它们定位于髓鞘、轴突、生长锥、线粒体膜及其他细胞骨架结构,表明CCI后存在突触重塑、轴突侧支芽生和代谢适应。此外,分子功能(MF)术语强调了钙离子结合和S100蛋白结合,支持其在Ca²⁺瞬变、激酶信号传导和突触前活动中的作用(图6c)。值得注意的是,S100蛋白在提议的疼痛神经通路的三个重要组织——坐骨神经(SN)、脊髓(SC)和眶额皮层(OFC)中均升高,提示疼痛神经通路中协调的可塑性和炎症反应。在我们的数据集中,我们还观察到OFC中调控多巴胺(DA)分泌的蛋白存在上调。相比之下,OFC中下调的DEPs富集于如小GTP酶介导信号转导的负向调节、有丝分裂细胞周期阶段转换的调节,以及吞噬作用和对抗细菌的防御反应等生物过程(图6d)。其相关的细胞组分包括细胞核、吞噬杯、细胞质核糖体和嗜天青颗粒。OFC中下调DEPs最显著的分子功能是视黄酸受体(RAR)结合。
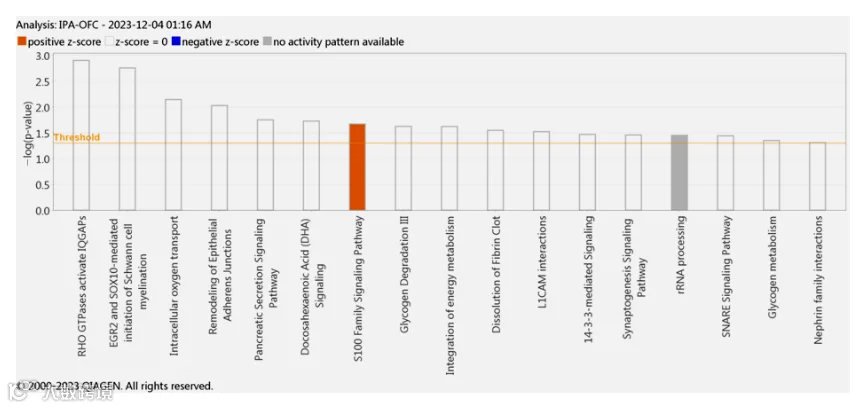
13.OFC蛋白质组的创新途径分析(IPA)
OFC蛋白质组的IPA分析识别出若干对NP病理生理学至关重要的经典通路和相互作用网络。根据z评分排名的前五条经典通路如下:RHO GTP酶激活IQGAP、ERG2和SOX10介导的施万细胞髓鞘形成启动、细胞内氧运输、上皮黏附连接重塑,以及胰腺分泌信号通路(表6)。这些通路提示损伤后SN中的肌动蛋白细胞骨架重塑,导致IQGAP与RHO GTP酶相互作用,从而增强TRPA1的膜转运并促进冷觉和机械性高敏感性。此外,OFC中的差异表达蛋白(DEPs)还参与糖原降解、二十二碳六烯酸信号传导及L1CAM相互作用。有趣的是,我们还发现OFC中表达的蛋白质激活了S100信号通路,这与其他被认为具有痛觉敏化作用的神经轴区域中观察到的结果类似(图7)。更重要的是,IPA识别出七个功能网络,其中包含我们数据集中的DEPs,这些网络可用于预测NP发病机制中的分子机制(表S4)。IPA所提出的最重要的网络在神经系统疾病、生物体损伤和异常以及骨骼和肌肉疾病中表现突出。因此,该网络最适用于NP病理生理学,NP是一种由神经系统损伤引起的神经性疾病,导致运动功能障碍。该网络富含与NP相关的蛋白质,如BDNF、CD200、CD388、CHRNA7、CHRNB2、EXTL2以及谷氨酸离子型受体亚单位(GRIN1、GRIN2A、GRIN2B、GRIN3A)(图S10),这些蛋白质在突触可塑性、免疫调节和兴奋性神经传递中起核心作用——这些都是NP发生和发展所必需的。
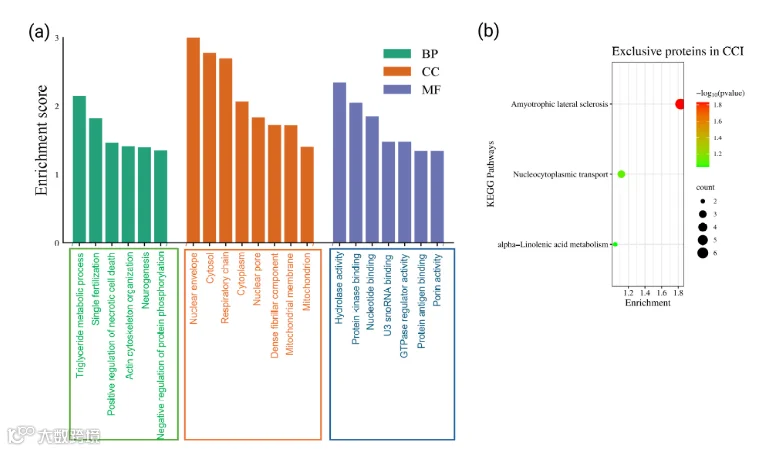
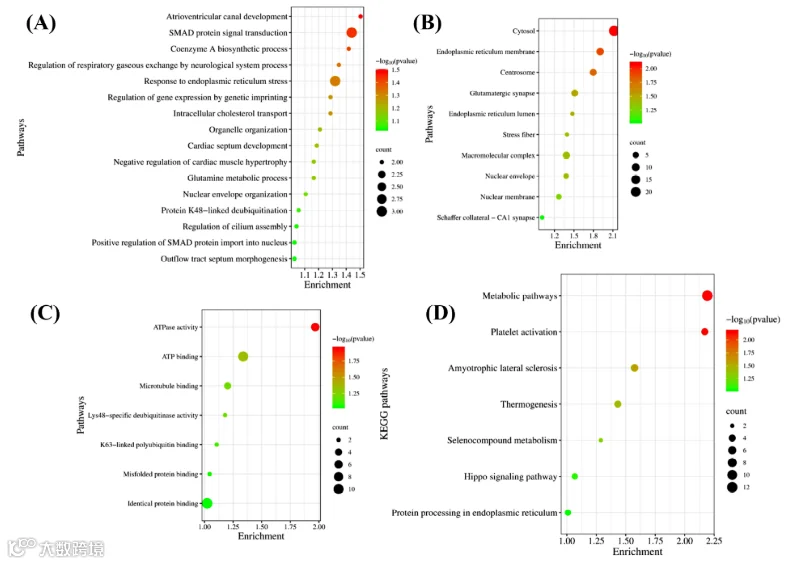
14.CCI组眶额叶皮质中特有表达蛋白的基因本体论和通路分析
在坐骨神经结扎(CCI)术后14天对大鼠眶额皮层(OFC)进行蛋白质组学分析,共鉴定出4600种蛋白质,其中83种仅在神经病理性疼痛(NP)动物中表达。在最丰富的CCI特有蛋白中包括ATP合酶、犬尿氨酸-草酰乙酸转氨酶3(KAT3)、周边素、S100-A11、基底细胞黏附分子、磷酸泛酰半胱氨酸脱羧酶、乙酰辅酶A转运蛋白SLC33A1、花生四烯酸15-脂氧合酶以及一个类似复合物III的蛋白。它们的选择性出现支持了这些蛋白可能参与NP的发生与进展的假说。因此,为了阐明这些CCI特有蛋白的作用,我们进行了GO和通路富集分析(图8)。这些蛋白显著富集的生物过程(BP)包括谷氨酰胺代谢过程、内质网应激反应、辅酶A生物合成过程、SMAD蛋白信号传导、细胞内胆固醇运输以及蛋白K48连接去泛素化(图8a)。同样,这些蛋白显著富集的细胞组分(CC)主要位于内质网膜、谷氨酸能突触、核膜、中心体、细胞质及应激纤维(图8b)。此外,它们的功能分子(MF)涵盖ATP酶活性、微管结合、Lys48特异性去泛素化酶活性以及K63连接多泛素结合(图8c)。此外,与这些特有蛋白相关的潜在且统计学显著的KEGG通路涉及代谢通路、血小板活化、内质网中的蛋白质加工以及Hippo信号通路(图8d)。
15.在提议的神经轴中周围神经系统与中枢神经系统之间的比较规范通路分析
周围神经损伤研究主要集中在周围神经和施万细胞(SC),而脊上区域在疼痛感知中的作用则相对研究较少。在我们的研究中,我们设计了一个框架来探索所提出的疼痛神经通路,连接外周神经系统(PNS)与中枢神经系统(CNS),以调查坐骨神经结扎(CCI)后背根神经节(SN)引发的系统性改变,这些改变有助于神经病理性疼痛(NP)的发生与发展。在这条通路上进行比较性的经典通路分析揭示了多个常见生化通路的富集,突显了疼痛处理在外周与中枢部位之间的分子交互作用(图9)。炎症级联反应在整个神经通路上尤为显著。中性粒细胞胞外陷阱形成、中性粒细胞脱颗粒以及S100家族信号传导是在SN、SC和额眶皮层(OFC)中富集最显著的通路之一(表7)。同时,在这三个区域中均观察到SNARE介导的胞吐作用、突触发生及NMDA受体信号通路的富集,强调了它们在兴奋性神经传递和适应不良可塑性中的关键作用。线粒体功能障碍以及sirtuin通路的激活也被观察到,提示氧化磷酸化和细胞能量稳态存在系统性损害。相比之下,内源性镇痛机制的上调,包括催产素和血清素受体信号传导,在脊上区域尤为明显,表明对慢性疼痛的代偿性反应。
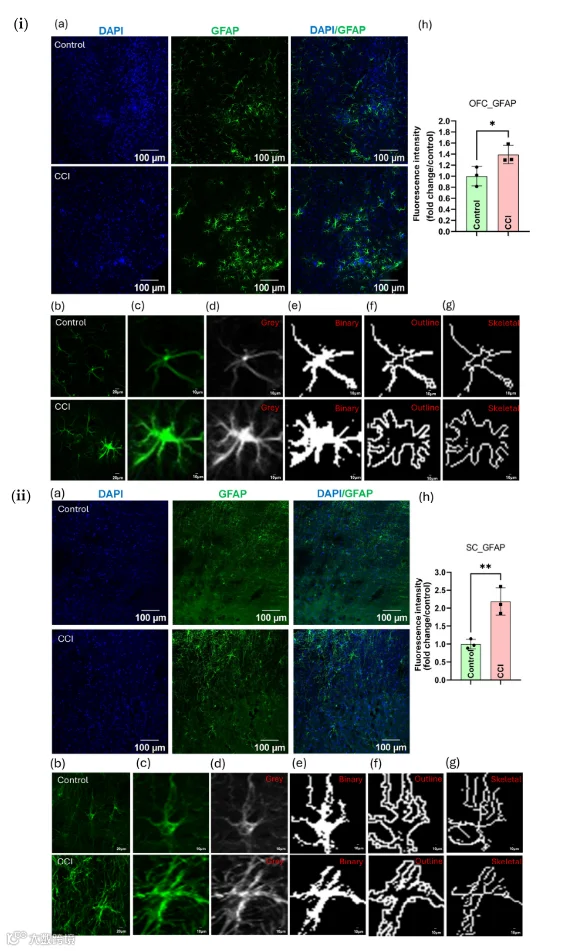
16.CCI诱导胶质细胞活化和S100蛋白上调及其在痛觉传导通路上相关的神经炎症
免疫荧光分析显示,在CCI术后沿拟议的伤害性神经轴路径上胶质细胞显著激活。具体而言,与对照组相比,CCI大鼠的OFC、SC和SN区域中GFAP阳性细胞数量明显增加(p < 0.05)(图10和S11)。更高倍数的图像显示,CCI动物中的GFAP阳性星形胶质细胞呈现肥大形态,其过程变粗、分支点增多、伸长及树突复杂性增强——这通过OFC和SC区域的二值化图、轮廓图和骨架图得到确认(图10i、ii分别)。
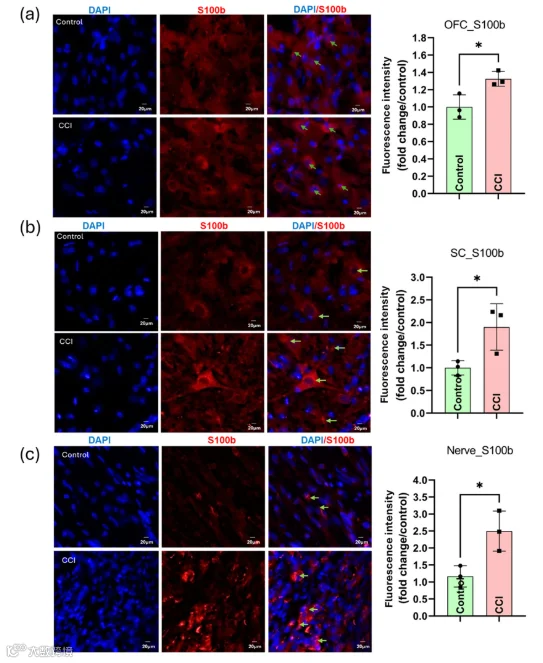
同样地,OFC和SC中均观察到小胶质细胞激活,表现为Iba-1表达增加以及胞体增大和突起收缩等形态学改变(图S12)。结构分析证实,CCI大鼠的小胶质细胞复杂性增加且分支减少(图S12c,f)。此外,周围神经损伤后,外周神经系统(PNS)和中枢神经系统(CNS)中促炎介质(如IL-6、TNF-α)水平升高,如图S13所示。此外,损伤相关分子模式(DAMP)S100A8在OFC、SC和SN中显著上调(p < 0.05;图S14)。另外,S100B在CCI后所有三个区域的表达均显著升高,如合并的DAPI/S100B图像和荧光定量所示(图11),支持系统性神经炎症反应(p < 0.05)。
17.外周神经的CCI增强中枢神经系统的多巴胺能信号传导
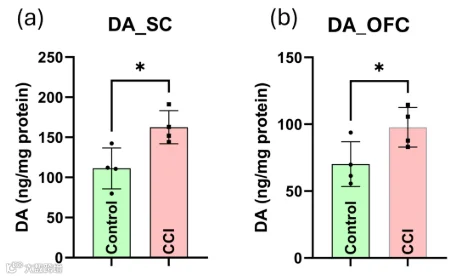
在CCI后,与对照大鼠相比,眶额皮层(OFC)和脊髓(SC)中的多巴胺水平显著升高(图12a,b),提示可能存在一种神经化学适应,该适应可能与胶质细胞介导的神经炎症反应相交集(p < 0.05)。此外,CCI模型中观察到的多巴胺升高可能代表一种不良适应性升高,从而促进神经敏化,也可能是一种稳态反应,试图激活内源性镇痛通路。
图12
更多结果和补充图表:doi:10.3390/cells15030290
长按二维码关注我们,用最短的时间和最高的效率学习更多数据分析方法!
扫描上方二维码或登录平台官网后添加CNSknowall客服微信咨询!官网地址:
https://cnsknowall.com
CNSknowall:24年最新问世的遥遥领先的科研数据(0代码生信+统计学)分析平台,同时含有机制图模块+汉化版Pubmed融合Deepseek高效筛选目标文献+SCI文献例句/语料检索模块+OPenAI官方GPT接口,>500款CNS级别图表皆可一秒内一键出图,登录即秒变数据分析大神,体验前所未有的便捷数据分析之旅,开启科研天骄之路!
可向下滑动批阅!