ARID1A是SWI/SNF染色质重塑复合物的关键组分,在约11.5%的黑色素瘤病例中发生突变。全基因组CRISPR筛选已将其与BRAF/MEK抑制剂耐药性相关联。然而,ARID1A突变的临床意义,特别是其如何协调早期信号适应与长期获得性耐药,以及如何影响肿瘤-免疫互作,仍不清楚。本研究通过整合多组学因子分析和网络传播方法,比较了药物敏感的ARID1A野生型与耐药的ARID1A敲除黑色素瘤细胞系对BRAF和MEK抑制剂的早期信号响应。旨在系统性地揭示ARID1A缺失所驱动的转录重编程、信号网络重塑及其在靶向治疗耐药和免疫逃逸中的作用。
今天给大家解读一篇2月发表在《Molecular Systems Biology》上的题目为“Integrative multi-omics defines melanoma drug response networks and ARID1A-dependent resistance mechanisms.”的文章。本研究针对黑色素瘤中BRAF/MAPK抑制剂耐药的挑战,通过分析BRAFV600E敏感细胞系和ARID1A敲除耐药细胞系,结合多组学方法,揭示了ARID1A缺失导致的信号通路重编程、免疫逃逸机制,并识别出关键耐药节点。(请持续关注我们,每天为您解读最新见刊的文献!)想薅生信资料羊毛?直接在对话框回复 “资料”,免费领取干货大礼包!
不想做实验,没数据,还想要快速发表文章,没问题的!公共数据库就是我们的数据宝藏!没思路不用担心,作为专业的生信团队,我们很乐意为你们效劳,提供研究路线设计和数据挖掘分析,扫码联系我们吧!


团队成员合影(位于上海陆家嘴中心,可随时预约参观)
题目:《整合多组学定义黑色素瘤药物反应网络和ARID1A依赖性耐药机制》Integrative multi-omics defines melanoma drug response networks and ARID1A-dependent resistance mechanisms
发表期刊: Molecular Systems Biology
影响因子:7.7
研究背景:
对BRAF/MAPK抑制剂的耐药是黑色素瘤治疗中的重大挑战,这一挑战由允许肿瘤细胞逃避治疗的适应性和获得性机制所驱动。ARID1A在黑色素瘤中频繁突变,与耐药和免疫逃逸有关。
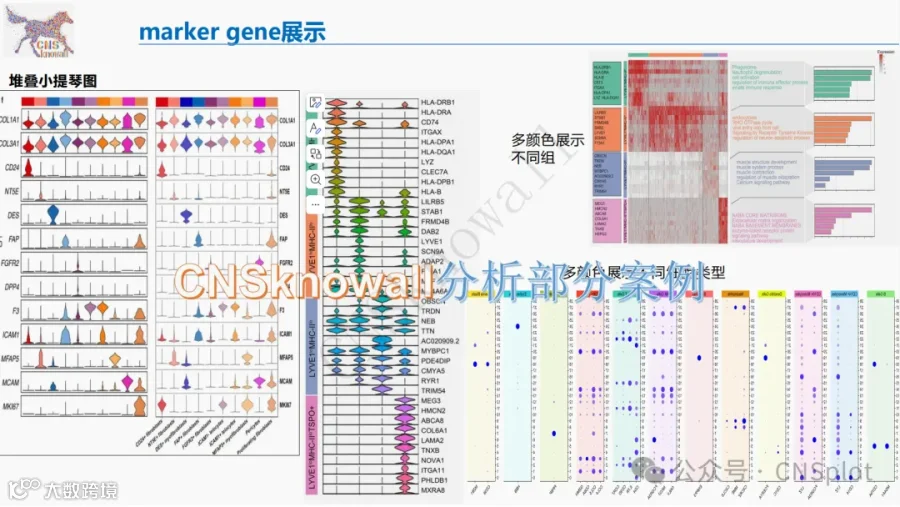
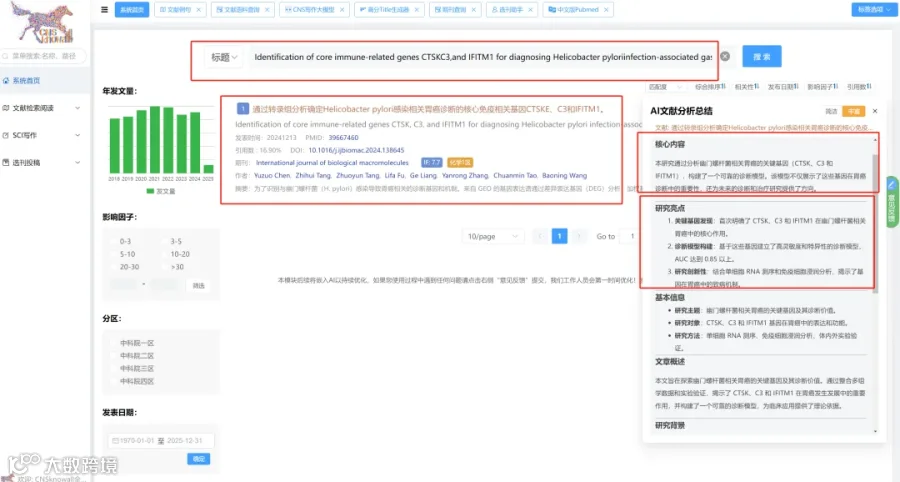
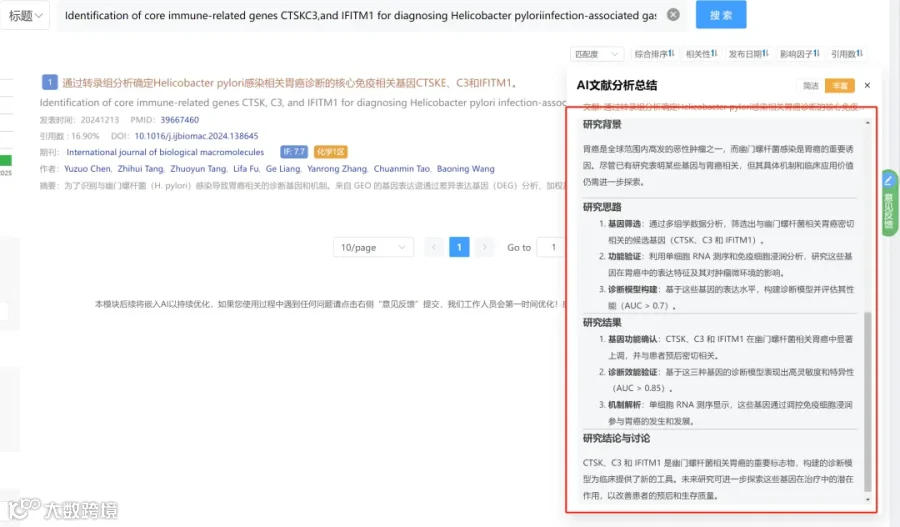
CNSknowall 平台 Pubmed+AI 快速提炼全文要点
研究思路:
在BRAFV600E敏感的黑色素瘤细胞系和药物耐药的ARID1A敲除(KO)衍生物中,探索对BRAF和MAPK抑制剂的早期信号响应。通过创新的系统生物学方法整合多组学数据集,识别关键耐药机制。
研究亮点:
-
揭示了ARID1A缺失如何驱动BRAF/MAPK抑制剂耐药。
-
识别出关键耐药节点(PRKD1、JUN、NCK1),为治疗策略提供潜在靶点。
-
-
发现ARID1A敲除影响HLA表达和细胞外基质,可能限制免疫疗法疗效。
研究结果:
-
ARID1A-KO细胞表现出转录重编程,在治疗后维持MAPK1/3和JNK活性,抑制PRKD1激活,增加JUN活性,并通过升高的RTKs(如EGFR、ROS1)和Ephrin受体活性扰乱PKC动态。
-
ARID1A-KO还减少了HLA相关蛋白的表达并增强了细胞外基质成分,可能限制免疫浸润和免疫疗法疗效。
-
多组学分析揭示PRKD1、JUN和NCK1作为关键耐药节点。
研究总结:
本研究通过整合多组学分析,定义了黑色素瘤药物反应网络和ARID1A依赖性耐药机制。关键耐药节点(PRKD1、JUN、NCK1)为对抗黑色素瘤耐药的潜在治疗策略提供了靶点。ARID1A缺失导致的免疫逃逸特征可能影响免疫疗法效果,这为进一步研究耐药调控机制和治疗干预方向提供了依据。
结果译文:
1.多组学数据整合识别与药物反应和ARID1A KO相关的分子特征
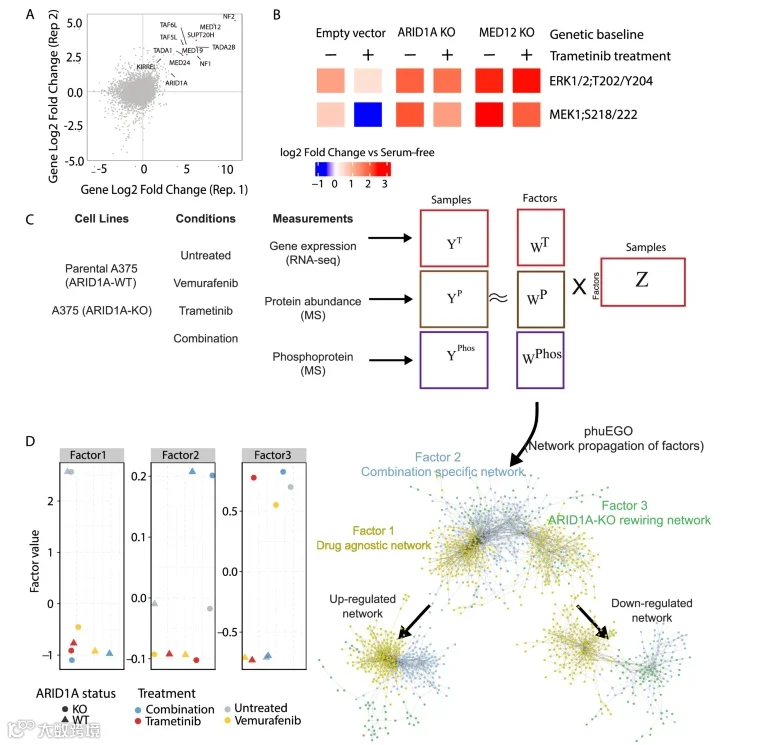
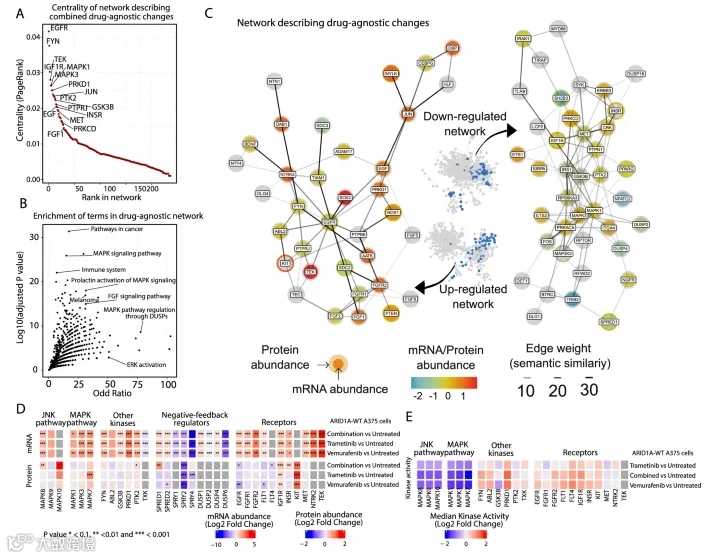
ARID1A此前在全基因组筛选中被确定为MAPK抑制剂(selumetinib)和BRAF抑制剂(vemurafenib)的耐药因子。为了评估ARID1A是否广泛作用于整个MAPK通路,而非介导对单一药物的特异性耐药,我们在A375细胞中使用了另一种MEK抑制剂trametinib进行了两次额外的筛选。这些筛选再次鉴定出ARID1A,以及已确定的耐药基因(如NF1/2、KIRREL、MED12、TAF5/6L),进一步支持了其在BRAFV600E黑色素瘤耐药中的作用。(图1A;数据集EV1)。用sgRNA靶向ARID1A或MED12(一个先前已被表征的vemurafenib耐药基因),随后用trametinib处理,导致pMEK和pERK磷酸化持续存在,表明细胞对MAPK抑制无响应(图1B)。
为了研究药物敏感与耐药ARID1A敲除细胞在信号传导和基因调控上的差异,我们使用了早期传代的ARID1A-WT A375亲本细胞和匹配的ARID1A-KO细胞系,后者对vemurafenib和trametinib表现出增强的耐药性。为了表征耐药相关的网络重塑,我们收集了来自亲本和ARID1A-KO细胞在有无药物处理6小时后的多组学数据(蛋白质组学、磷酸化蛋白质组学和转录组学);图1C)。选择此时间点是因为它能提供稳态测量,并通过初步的17重Luminex测定确认了MAPK抑制剂对磷酸化的抑制作用。
通过质谱分析,我们在整合不同实验批次后,定量了8139种蛋白质和3207个磷酸化位点。利用RNA-seq,我们定量了14,376个基因的转录水平。所有数据集均具有可重复性。在ARID1A-WT与KO实验间差异显著的715种蛋白质、372个磷酸化肽段和7557个基因中,仅有13个是所有数据集共有的,证明了不同组学层次的互补性。
我们应用多组学因子分析来无偏地识别驱动所有数据模态变异的潜在因子(图1C)。这揭示了三个关键因子:因子1和2捕获了对药物处理的适应性反应,因子3反映了由ARID1A缺失导致的持续性耐药。低维数据表示显示,与联合治疗相比,单一药物治疗在组学水平上的差异很小。方差分析表明,药物相关因子解释了蛋白质、磷酸化位点和mRNA水平的大部分变异。因子2与联合治疗相关,对转录组无影响,但解释了磷酸化蛋白质组和蛋白质组的显著方差。因子3与ARID1A KO相关,主要影响转录组和蛋白质组。
为了理解上述因子所代表的信号过程的功能含义,我们试图将鉴定出的基因置于其功能环境(即其相互作用网络)的背景中。为此,我们改进了phuEGO,一种基于网络传播的方法,用于从磷酸化蛋白质组数据中提取活性特征,并与MOFA的因子载荷结合使用。PhuEGO将网络传播与自我网络分解相结合,能够识别由与输入节点功能上和拓扑上最相似的节点组成的小型网络。这使得我们能够从因子特异性载荷中生成最小网络,这些网络覆盖了驱动药物反应和ARID1A KO差异的蛋白质,作为因子权重的函数。分别对正负权重进行处理后合并,产生三个网络。得到的正负权重网络分别代表与每个因子相关的上调或下调信号模块,从而捕获与药物反应或ARID1A缺失相关的对立功能程序。
图1
2.与治疗相关的药物非特异性变化涉及RTK和MAPK的负反馈
如上所述,因子1捕获了治疗非特异性变化,即无论BRAF、MEK1/2还是双重抑制均发生的改变。该网络的中心节点包括关键受体、激酶和转录因子,如EGF、FYN、IGF1R、TEK、MAPK1和MAPK3,以及PRKD1、JUN、PTK2和INSR(图2A;附录图S2A)。因子1相关网络的通路富集分析突显了与“黑色素瘤”和MAPK信号相关的术语,包括“DUSP对MAPK通路的调节”(图2B,C)。
与其它研究一致,我们在因子1的phuEGO衍生下调网络中观察到了负反馈调节因子。这与我们的原始组学数据相符,我们看到已知的MAPK负反馈调节因子丰度下降,这些因子被证明与MAPK3相互作用。在SPRY1/2/4和SPRED1/2中,我们检测到RNA丰度和蛋白质丰度均下降。这些RTK和MAPK信号负调节因子的减少似乎能解除下游抑制,导致生长因子信号增加,并减轻vemurafenib或trametinib对MAPK抑制的效果。
我们还在phuEGO衍生的上调网络中发现了几个受体酪氨酸激酶和其他激酶,包括EGFR、ROS1、FGFR1和2、KIT、FYN等,表明药物处理后许多通路存在广泛的信号调控和重 wiring。蛋白质/转录组丰度与激酶活性相关性不佳,我们只发现少数磷酸化位点被调节。因此,为了研究药物处理后观察到的信号调控,我们还使用PamChip技术收集了功能性激酶组学数据,该技术通过一系列固定的靶标肽阵列,提供细胞裂解液中多重激酶活性的估计。该数据集中激酶活性的变化幅度与我们网络中激酶的中心性相关性良好,其中MAPK1和PRKD1是我们网络中定量到的最中心的激酶。其中,我们发现药物处理后MAP激酶活性显著降低,尽管这并未反映在它们各自丰度的变化上。我们还发现了其他几种激活的激酶,包括PRKD1、FYN和IGFR1,这些在我们的网络中显示为中心节点,以及ABL2、FLT1和FGFR2。EGFR仅表现出非常小的活性增加,这与它在转录组和蛋白质组丰度上的减少形成对比。总之,这些结果表明药物扰动后存在RTK驱动信号的重 wiring。
图2
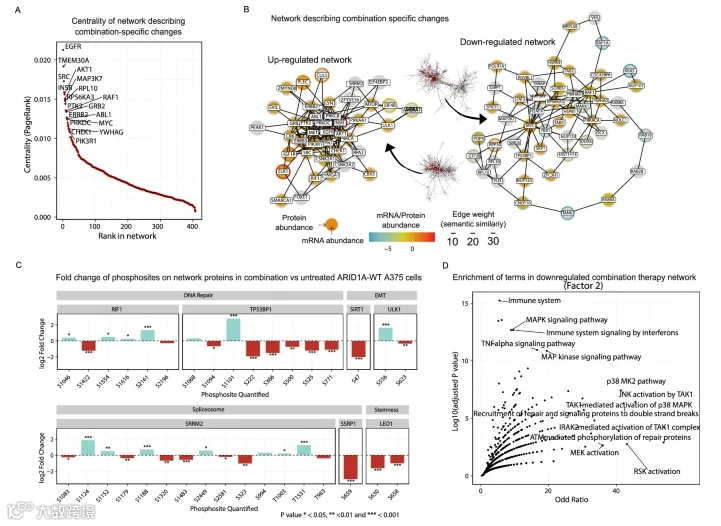
3.联合疗法诱发与DNA损伤修复相关的磷酸化模式
因子2代表了与联合用药反应相关的改变(图1D),并且主要来自磷酸化蛋白质组数据。同时靶向BRAF和MEK的联合用药是目前转移性和不可切除黑色素瘤患者的标准治疗方案,并已在患者中显示出更长的疾病缓解期。我们的功能性激酶组学数据表明,在我们的实验中,联合治疗与vemurafenib单药治疗对MAP激酶的影响几乎没有差异。我们也没有观察到该细胞系联合治疗相比单药治疗的杀伤效率差异。因此,我们决定聚焦于此因子,以阐明在此水平上观察到的磷酸化蛋白质组差异。
将phuEGO应用于因子2的权重,绘制了受联合治疗影响的独特蛋白质网络。我们检测到跨膜受体,特别是ERBB2、INSR和EGFR,在网络中处于中心位置,并被差异磷酸化的蛋白质包围。参与细胞迁移的TMEM30A是第二中心节点,其次是SRC。MAPK之外的替代生长调节因子AKT1也具有很高的中心性。相应的联合治疗特异性下调网络以转录因子MYC和DNA损伤反应蛋白TP53BP1为中心。信号转导因子RAF1和KRAS也存在。同时观察到MAP3K7,由于联合抑制MAP2K1和BRAF后其磷酸化减少而具有负权重。
尽管磷酸化蛋白质组是该因子的主要驱动因素,但与单药扰动相比,功能性激酶组学数据中的激酶活性变化很小。观察驱动因子2方差的磷酸化位点,我们发现几个参与DNA修复相关功能的磷蛋白。例如,TP53BP1在丝氨酸残基1101上高度磷酸化,而在联合治疗后,从残基222到771的多个丝氨酸残基上的磷酸化水平显著降低。TP53BP1能够促进DNA双链断裂的NHEJ修复。已知丝氨酸残基1101的磷酸化与电离辐射诱导的DNA损伤相关。RIF1是TP53BP1的关键调节因子,其在联合治疗后也通过磷酸化被显著调节。虽然这些位点没有功能注释,但联合治疗中下调的RIF1-S2196非常接近已知抑制该蛋白功能的S2205,并且预测可被JNK1、3或P38δ或γ等激酶磷酸化,所有这些激酶在我们的功能性激酶组学数据中相比无药物处理时均下调。已知参与DNA修复过程的SSRP1在S659位点磷酸化减少。已知被ATM磷酸化并能诱导自噬的ULK1-S556,在联合治疗相比所有其他条件下表现出强烈的上调。SIRT1-S47不是驱动因子2的磷酸化位点,但它是联合治疗相比未处理对照显著下调的肽段之一,已知通过自噬降解E-钙粘蛋白促进上皮-间充质转化。参与维持胚胎干细胞多能性的LEO1-S630也下调。最后,我们观察到剪接体组分SRRM2在多个磷酸化位点上的磷酸化调控。
放大视野观察该网络所涉及的过程,我们对上下调网络进行了功能富集分析。上调网络显示出RTK驱动通路的富集,以及PI3K/Akt和mTOR信号传导。下调网络反映了预期的MAPK通路抑制,并包含与DNA损伤修复和免疫信号相关的术语。
图3
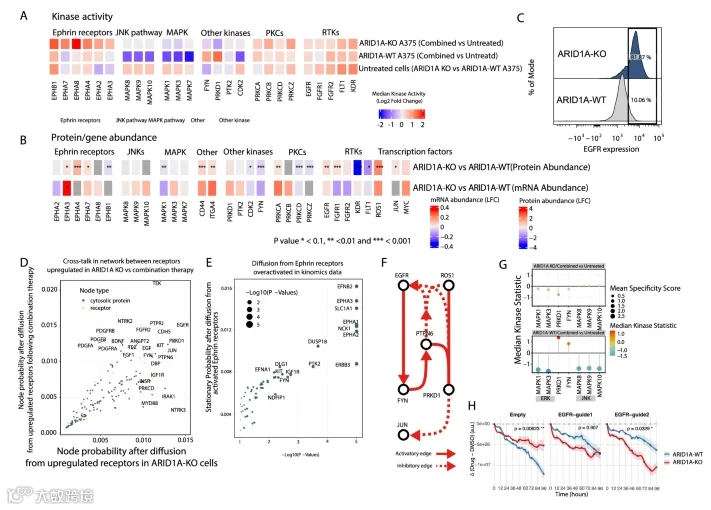
4.ARID1A KO后细胞的 basal 转录状态影响对药物处理的反应
尽管ARID1A-KO细胞对MEK/BRAF抑制具有耐药性(图4A),但其转录组和蛋白质组对药物的反应与亲本细胞高度相似(图1D和图EV4A,B)。这体现在ARID1A-KO与ARID1A-WT细胞在药物处理后的差异丰度上,转录组和蛋白质组反应均具有高度相关性。然而,激酶活性测定和磷酸化蛋白质组学揭示了信号行为的显著差异。例如,与亲本A375相比,ARID1A-KO A375细胞中MAPK1/3对药物处理的敏感性降低,而Ephrin受体信号则升高(图4A)。
为解释这一现象,我们聚焦于两种细胞系间的差异,并发现了显著不同的 basal 状态。具体而言,ARID1A缺失作为转录重编程因子,重塑了初始基因和蛋白质表达 landscape,从而重 wiring 信号传导并改变对药物扰动的响应。与亲本细胞相比,ARID1A-KO细胞显示出几种受药物疗法影响的受体丰度增加,包括ROS1、ITGA4、EGFR和CD44(图4B)。特别是EGFR,在ARID1A-KO系中的膜表达相比亲本系急剧增加,这与耐药黑色素瘤细胞中受体内吞机制受损一致(图4C)。致癌转录因子如JUN和MYC及其调节子也上调(图4B和图EV4D),表明ARID1A缺失使细胞为增强的致癌信号和药物耐药做好了准备。
为了探究这些 basal 变化的功能效应及其如何影响治疗反应,我们模拟了ARID1A KO诱导的受体变化与药物诱导的受体改变之间的信号 crosstalk。为此,我们首先整合了捕获药物非特异性反应信号的网络与ARID1A KO特异性效应的网络,生成了与两者相关的过程的组合图谱。该网络内通路的富集揭示了MAPK信号、免疫系统通路、黑色素瘤、粘着斑和FGF信号中的强烈信号。接下来,使用马尔可夫随机游走方法,我们分别从由ARID1A KO上调的受体和由药物反应诱导的受体传播信号,以识别 crosstalk 的点。该分析突显了两种信号汇聚的共享枢纽,包括JUN、PRKD1、FYN、PTPN6和PRKCD(图4D)。我们接下来检查了Ephrin受体家族,该家族在ARID1A-KO而非亲本细胞中,在药物处理后强烈激活(图4A,E;附录图S2C)。从这些受体进行的网络传播识别了额外的参与者,如NCK1、DUSP18、PTK2、FYN和DAPK,作为潜在的介导者(图4E)。
比较ARID1A-KO的 basal 蛋白质重编程与药物诱导的丰度变化进一步阐明了这些效应。在ARID1A-KO细胞中,NCK1丰度对药物处理无响应,而DUSP18获得了响应性。尽管FYN和DAPK1在亲本和KO细胞中显示出相似的治疗反应,但它们的 basal 丰度发生了改变,从而改变了通过这些节点的有效信号流量(附录图S4C)。
为了识别这种重 wiring 通讯的关键路径,我们进行了最大流网络分析,以追踪向下游效应器的最可能信号路径。这揭示了从EGFR和ROS1通过中间体(包括PRKD1、FYN和PTPN6)汇聚到JUN的流动(图4F),这与我们在马尔可夫传播分析中它们被鉴定为 crosstalk 枢纽的结果一致。功能性激酶测定证实,与亲本细胞不同,在ARID1A-KO细胞中,FYN和PRKD1均表现出药物诱导的激活减弱(图4G),证明这些节点在功能上被重 wiring,并且不再对MEK/BRAF抑制做出动态响应。
为了直接测试对上游受体信号的预测依赖性,我们在ARID1A-KO和亲本ARID1A-WT系中使用两种不同的gRNA靶向EGFR,并通过IncuCyte成像系统在96小时内测量联合药物处理后的细胞活力。EGFR缺失消除了ARID1A-KO细胞的耐药表型,并使细胞死亡水平超过亲本细胞中观察到的水平(图4H),证实了EGFR驱动的信号是ARID1A介导的耐药性的关键决定因素。
总之,这些结果表明ARID1A缺失重塑了受体丰度、膜信号组织和转录活性。尽管全局的转录组和蛋白质组药物反应在基因型间看似相似,但改变的 basal 信号拓扑结构将来自EGFR和ROS1的流通过重 wiring 的FYN-PRKD1-JUN轴重新定向,解释了ARID1A-KO细胞独特的药物反
图4
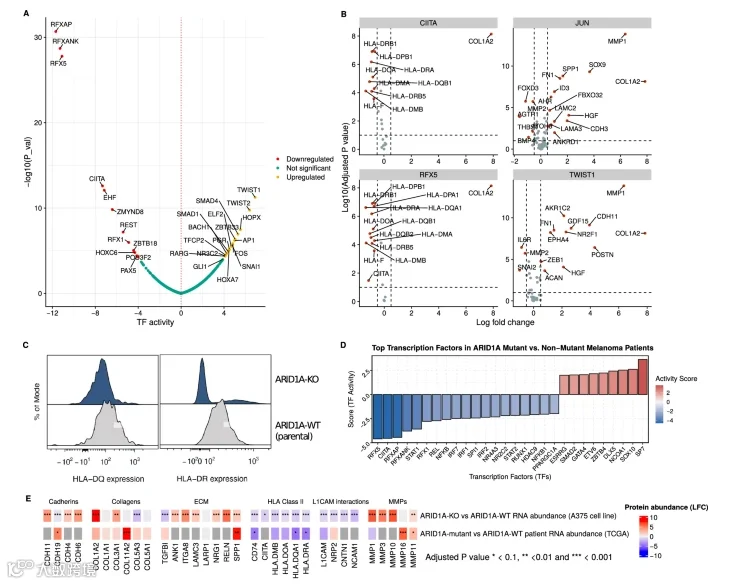
5.ARID1A KO在体外和体内环境下均抑制HLA蛋白表达
为了评估ARID1A KO如何影响药物处理下的转录调控,我们分析了药物处理后的转录因子活性。我们观察到ARID1A KO后JUN和MYC活性的 basal 水平增加,与我们之前的发现一致。值得注意的是,我们也观察到调节MHC II类基因转录的RFX5、RFXAP、RFXANK和NFYC的调节子活性显著下降(图5A)。这导致MHC II类基因和CD74的表达降低,而被RFX5抑制的基因则增加(图5B)。
虽然MHC II类分子通常表达于专职抗原呈递细胞,但A375细胞也表达它们。为了证实我们的发现,我们发现ARID1A-KO细胞表面HLA-DQ和HLA-DR的表达显著降低(图5C)。此外,上调的转录因子包括ECM重塑的调节因子,如TWIST和SMAD家族(图5A,B)。
利用TCGA数据,我们调查了这些模式是否出现在黑色素瘤患者中。我们将472名患者分为ARID1A受影响组和未受影响组。ARID1A受影响的患者显示出MHC II类调节因子RFXAP、RFX5和CIITA的预测活性显著降低。此外,胶原蛋白和金属蛋白的调节因子SP7显著上调。ARID1A受影响的患者和KO细胞均表现出胶原蛋白和层粘连蛋白表达增加,可能促成ECM硬度,以及HLA蛋白水平降低(图5E)。
图5
更多结果和补充图表:doi:10.1038/s44320-025-00183-5
长按二维码关注我们,用最短的时间和最高的效率学习更多数据分析方法!
扫描上方二维码或登录平台官网后添加CNSknowall客服微信咨询!官网地址:
https://cnsknowall.com
CNSknowall:24年最新问世的遥遥领先的科研数据(0代码生信+统计学)分析平台,同时含有机制图模块+汉化版Pubmed融合Deepseek高效筛选目标文献+SCI文献例句/语料检索模块+OPenAI官方GPT接口,>500款CNS级别图表皆可一秒内一键出图,登录即秒变数据分析大神,体验前所未有的便捷数据分析之旅,开启科研天骄之路!
可向下滑动批阅!