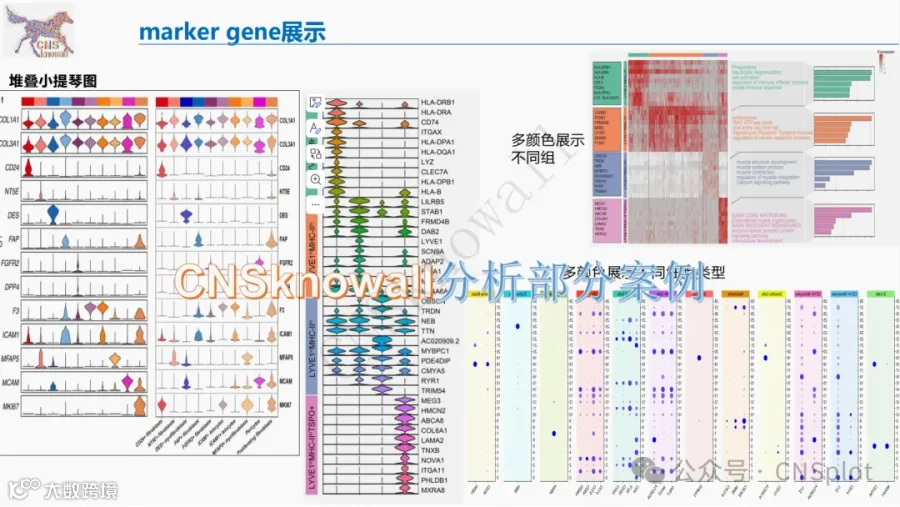
小胶质细胞能否被外周单核细胞替代?替代后它们会变成什么?本研究通过单细胞转录组学、ATAC‑seq 与 CUT&Tag 表观组学 等多组学技术,揭示了在反复小胶质细胞清除后,外周单核细胞可大量植入脑实质,获得与卵黄囊来源小胶质细胞截然不同的转录组与表观组身份。脑内植入的单核细胞衍生巨噬细胞(MDM)不仅呈现边界相关巨噬细胞样特征,还会短暂表达 CD206,并受 IL‑34 与 CCR2 调控。更令人意外的是,血液与颅骨骨髓来源的单核细胞植入脑内后呈现不同的分子状态,且 MDM 植入会加重脱髓鞘病理。该研究为理解脑内巨噬细胞的来源、身份与功能提供了全新多组学视角。
今天给大家解读一篇3月发表在《Neuron》上的题目为“Brain-engrafted monocyte-derived macrophages from blood and skull-bone marrow exhibit distinct properties.”的文章。本研究旨在厘清脑实质巨噬细胞在稳态、更新及病理过程中的组成与功能。通过综合运用多种实验技术,研究证实了脑内存在由单核细胞植入形成的巨噬细胞(MDMs),其尽管与卵黄囊来源的小胶质细胞(YSMs)共享微环境,但拥有截然不同的分子特征。研究进一步阐明了调控MDM植入的关键因子,并首次揭示颅骨骨髓与血液共同作为这些细胞的来源,贡献不同状态的MDMs。功能实验表明,MDMs在脱髓鞘病理中具有重要作用。(请持续关注我们,每天为您解读最新见刊的文献!)想薅生信资料羊毛?直接在对话框回复 “资料”,免费领取干货大礼包!
不想做实验,没数据,还想要快速发表文章,没问题的!公共数据库就是我们的数据宝藏!没思路不用担心,作为专业的生信团队,我们很乐意为你们效劳,提供研究路线设计和数据挖掘分析,扫码联系我们吧!


团队成员合影(位于上海陆家嘴中心,可随时预约参观)
题目:《来自血液和颅骨骨髓的脑移植单核细胞来源巨噬细胞表现出不同的特性》Brain-engrafted monocyte-derived macrophages from blood and skull-bone marrow exhibit distinct properties
发表期刊:Neuron
影响因子:15
研究背景:
小胶质细胞被认为起源于卵黄囊祖细胞,并在整个生命过程中持续存在,几乎不依赖于成年期造血系统的补充。然而,脑内植入的单核细胞来源巨噬细胞(MDMs)是否存在于稳态和更新过程中,以及它们相对于卵黄囊来源的小胶质细胞(YSMs)的功能如何,仍未明确。
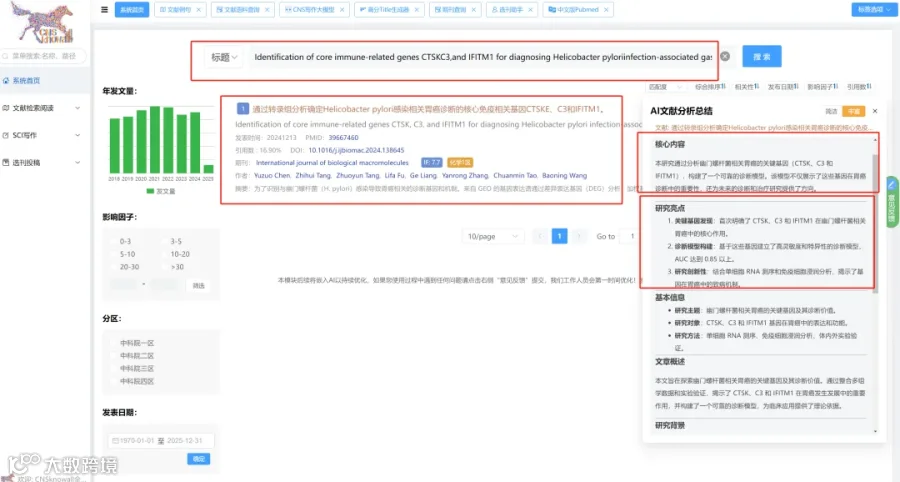
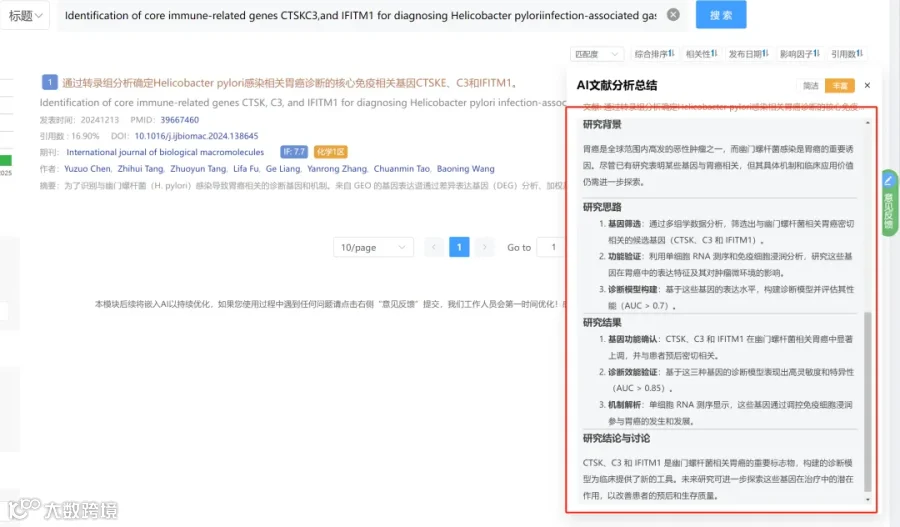
CNSknowall 平台 Pubmed+AI 快速提炼全文要点
研究思路:
研究者结合了谱系追踪(以确定细胞起源)、药理学小胶质细胞耗竭(以创造植入机会并研究更新过程)以及多组学分析(转录组和表观遗传,以定义分子特征)来系统探究脑实质巨噬细胞。同时,通过联体共生和颅骨瓣移植实验来追溯MDMs的具体解剖学来源,并通过构建脱髓鞘疾病模型来评估其功能。
研究亮点:
-
证实了在稳态和更新过程中,脑内存在植入的单核细胞来源巨噬细胞(MDMs)。
-
揭示了MDMs与经典卵黄囊来源小胶质细胞(YSMs)在转录组和表观遗传景观上的根本差异。
-
发现MDMs短暂表达CD206,类似于小胶质细胞前体的一个发育阶段。
-
明确了IL-34和CCR2信号调控MDM的植入与极化。
-
通过联体共生和颅骨瓣移植实验,证明血液和颅骨骨髓共同作为MDM的生态位,并产生具有起源偏向性的细胞状态。
-
研究结果:
- 身份差异
脑内植入的MDMs表现出与YSMs不同的转录和表观遗传景观。
- 发育特征
命运图谱显示,脑内植入的MDMs会短暂表达CD206,这模仿了小胶质细胞前体的一个发育阶段。
- 调控机制
MDM的植入和极化受到白介素(IL)-34和C-C趋化因子受体2(CCR2)的调控。
- 细胞来源
联体共生和颅骨瓣移植实验表明,血液和颅骨骨髓均能向脑实质供应MDMs,并产生具有起源偏向性状态的细胞。
- 病理功能
研究总结:
本研究系统性地定义了脑实质巨噬细胞在不同生理和病理条件下的组成。结论表明,成年脑实质中存在功能特化的单核细胞来源巨噬细胞(MDMs),它们与经典的小胶质细胞(YSMs)在起源、分子特征和功能上均存在区别。这些MDMs来源于循环血液和邻近的颅骨骨髓,其行为受特定的细胞因子和趋化因子受体信号通路调控。在疾病情境下(如脱髓鞘),MDMs的植入会加剧病理过程。该研究深化了对中枢神经系统巨噬细胞异质性及其在健康和疾病中作用的理解。
结果译文:
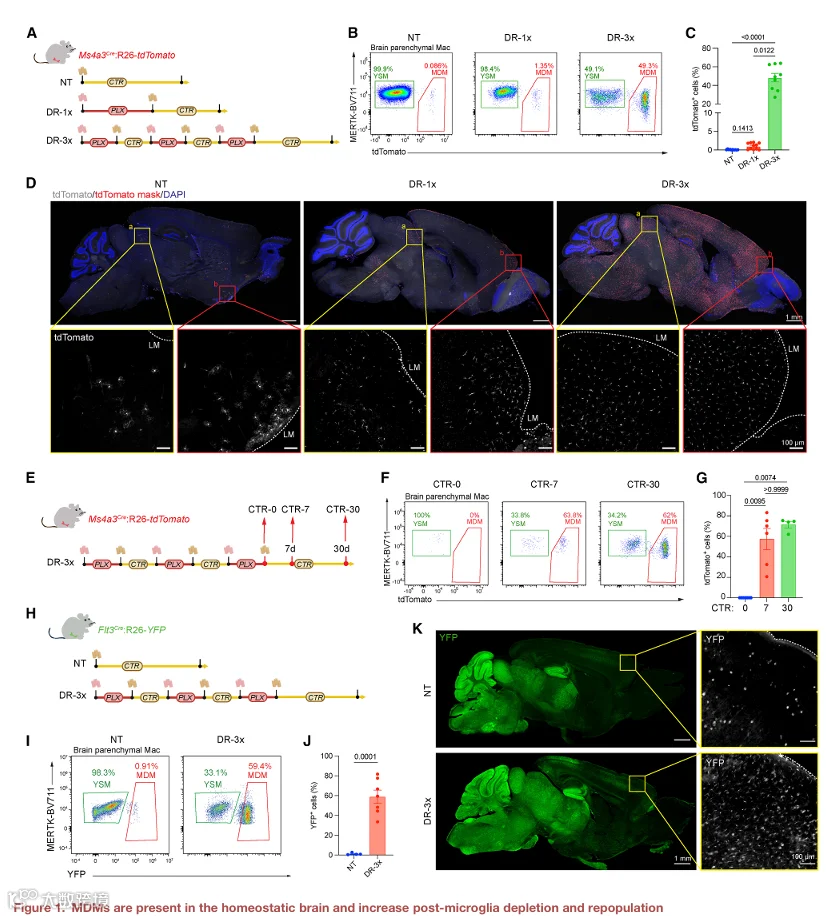
1.MDM 存在于稳态大脑中,并在小胶质细胞清除后增加
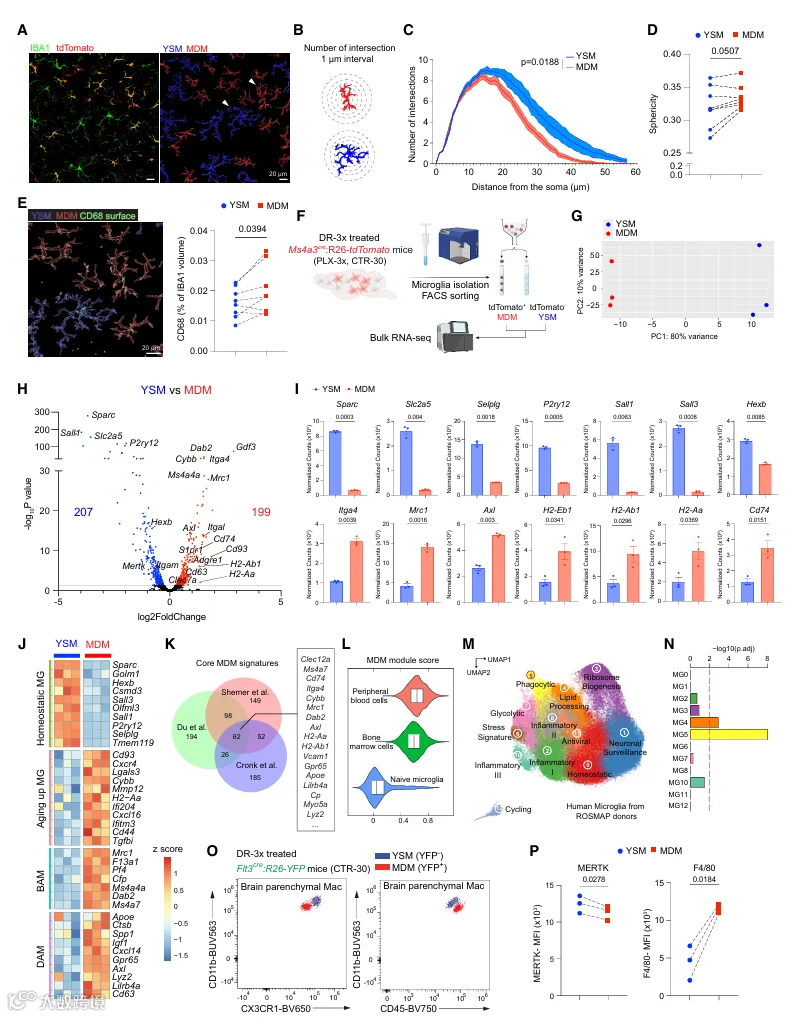
鉴于 Ms4a3 在粒细胞-单核细胞祖细胞及其单核细胞后代中选择性且瞬时表达,Ms4a3Cre;R26‑tdTomato 小鼠能够区分 MDM(tdTomato⁺)与 YSM(tdTomato⁻)(图 S1A)。为了研究小胶质细胞更新过程中脑内植入 MDM 的存在情况,我们给小鼠喂食 PLX5622(一种 CSF1R 的药理学抑制剂)以清除小胶质细胞。我们进行了单轮或三轮的清除-再群体化(DR‑1x 或 DR‑3x;图 1A)。已有研究表明,DR‑3x 会导致小胶质细胞长期缺失且重建速度减慢,但再群体化小胶质细胞的来源尚未明确。流式细胞术显示,未经处理小鼠的小胶质细胞群中 tdTomato⁺ MDM 极少(0.09% ± 0.03%)(图 1B 和 1C;门控策略见图 S1B)。DR‑1x 仅诱导 tdTomato⁺ MDM 轻度增加(1.00% ± 0.26%),而 DR‑3x 导致 tdTomato⁺ MDM 急剧扩增,约占所有脑实质巨噬细胞的一半(48.33% ± 4.51%)(图 1D 和 S1C)。为确定这种富集是否反映了 tdTomato⁺ 细胞对 PLX 的选择性抵抗,我们在最后一个 PLX 周期结束后、返回对照饮食前立即分析了大脑(CTR‑0)(图 1E‑1G)。此时,所有剩余的脑实质巨噬细胞均为 tdTomato⁺,表明 tdTomato⁺ 细胞对 PLX 并无优先抵抗性。相反,tdTomato⁺ 细胞的增加发生在 PLX 撤除后的再群体化阶段,很可能是由于外周细胞的招募。值得注意的是,在 PLX 撤除后 1 周(CTR‑7)即可检测到 tdTomato⁺ MDM,表明外周植入在再群体化早期就已开始(图 1E‑1G)。
脑内植入 MDM 的空间分布在不同的处理条件下有所不同(图 1D)。在未经处理的 Ms4a3Cre;R26‑tdTomato 小鼠中,罕见的 tdTomato⁺ MDM 定位于软脑膜附近,特别是上丘和腹侧纹状体,并呈现出高度分支的形态(图 1D)。tdTomato⁺ MDM 也呈 IBA1 阳性(图 S1C)。DR‑1x 使上丘和额叶皮层中的 tdTomato⁺ MDM 轻度增加,而 DR‑3x 导致其在大多数脑区广泛扩增,嗅球中的细胞相对较少(图 1D 和 S1D‑S1F)。DR‑3x 还产生了大量 IBA1⁺ MHC I 类高表达的实质巨噬细胞,特别是在嗅球中(图 S1D‑S1F),表明再群体化巨噬细胞的来源和表型存在区域异质性。
为了进一步验证我们的发现,我们采用了 Flt3Cre;R26‑YFP 谱系追踪模型。与 Ms4a3Cre 一样,Flt3Cre 标记来自成体骨髓的 MDM。此外,它还标记源自胎肝的 HSC 后裔(图 1H)。尽管该模型中 YFP 对神经元有背景标记,可能反映了神经元 Flt3 的表达(图 1I、S1G 和 S1H),但在未经处理的动物中,只有一小部分脑巨噬细胞为 YFP⁺(1.22% ± 0.52%),而 DR‑3x 使 YFP⁺ MDM 增加到脑巨噬细胞的 58.74% ± 6.71%(图 1I‑1K)。
2.脑内植入 MDM 获得与小胶质细胞不同的转录组和表观组身份
为阐明脑内植入 MDM 的分子身份,我们对从 DR‑3x 处理的 Ms4a3Cre;R26‑tdTomato 小鼠中分选的 tdTomato⁺ MDM 和 tdTomato⁻ YSM 进行了 bulk RNA‑seq(图 2A)。PCA 将 YSM 与 MDM 明显分开,表明两者具有不同的转录组谱(图 2B)。YSM 中上调的基因(例如 P2ry12、Tmem119、Sall1、Cx3cr1)与已报道的小胶质细胞身份基因一致(图 2C)。相比之下,MDM 中上调的基因(例如 Cd74、Mrc1、Axl、Itga4、Cxcl16)富集于吞噬体形成、细胞粘附、抗原加工提呈以及 IFN‑γ 应答等通路(图 2D 和 S2A)。MDM 还表达高水平的 Clec12a,这是一种单核细胞/巨噬细胞标志物,以及 Ccr2,这是单核细胞募集的趋化因子受体(图 2C)。为了评估这些转录组差异是否与形态学相关,我们对 DR‑3x 处理后的脑切片进行了成像分析。YSM 表现出典型的高度分支形态,而 MDM 则表现出圆形、阿米巴样形态,伴有较少的突起,且细胞体增大(图 2E 和 2F)。对 DR‑3x 处理小鼠分选的 MDM 进行透射电子显微镜分析进一步证实,MDM 的细胞质中电子致密颗粒和液泡数量增加(图 2G),并且包含髓鞘碎片样结构(图 2H)。
为了确定 MDM 获得的转录组身份是否受其植入的脑实质微环境的影响,我们将 DR‑3x 处理后分选的 MDM 与来自 Ms4a3Cre;R26‑tdTomato 小鼠脾脏的单核细胞(Spl Mo)以及从对侧半球硬脑膜中分离的硬脑膜巨噬细胞进行了比较(图 S2B)。PCA 显示 MDM 转录组介于脾脏单核细胞与硬脑膜巨噬细胞之间,表明脑实质微环境在塑造其身份方面发挥着作用(图 S2C)。然而,MDM 并未获得小胶质细胞样的转录组状态,这与先前的发现一致。我们进一步探究了 MDM 是否更接近边界相关巨噬细胞。在硬脑膜巨噬细胞中,我们观察到与 MDM 共享的 Mrc1、Cd74、Axl、Itga4 和 Cxcl16 上调,而 YSM 则富集 Tmem119、Sall1 和 P2ry12(图 S2D)。此外,MDM 和硬脑膜巨噬细胞在多种促炎和抗炎基因上表现出相似的表达模式(图 S2E)。在 MDM 转录谱中未检测到明显的性别依赖性差异(图 S2F‑S2G)。
为了定义跨植入范式的核心 MDM 基因特征,我们将来自 DR‑3x 数据集的 MDM 特征基因与两个独立的基于照射的骨髓移植数据集取交集。该分析确定了一组共 62 个共享基因——包括 Clec12a、Ms4a7、Cd74、Itga4、Mrc1、Dab2 和 Axl——我们将其定义为核心脑内植入 MDM 特征(图 2K;表 S1)。然而,我们注意到来自 DR‑3x 范式的 MDM 特征与来自 IR‑BMT 研究的特征重叠有限。为了区分范式特异性效应与批次效应,我们并行进行了 IR‑BMT 和 DR‑3x,并对两种条件下的 MDM 进行了谱分析(图 S2H 和 S2I)。PCA 和样本间距离分析显示,YSM 在范式间保持转录组相似性,而 DR‑3x 和 IR‑BMT 来源的 MDM 则表现出显著差异(图 S2J 和 S2K)。IR‑BMT 来源的 MDM 富集了与氧化磷酸化、IFN‑γ 应答和 TNF‑α 信号传导相关的基因(图 S2L 和 S2M)。一致地,与 DR‑3x 相比,IR‑BMT 脑裂解物中的 IFN‑γ 水平升高(图 S2N)。尽管存在这些实验差异,关注跨范式共享的基因能够定义稳健的、依赖于发育起源的核心 MDM 程序。
我们通过在一个已发表的小鼠单细胞 RNA‑seq 数据集(包含初始小胶质细胞以及源自骨髓或外周血的小胶质细胞样细胞)中进行模块评分,验证了这一核心 MDM 特征(图 2L)。随后,我们利用来自宗教秩序研究和记忆与衰老项目的人小胶质细胞图谱,探究人脑中是否存在类似群体(图 2M)。富集分析显示,吞噬性小胶质细胞簇 5 对核心 MDM 特征的富集最强(图 2N)。MG5 的特征是清道夫受体(如 MRC1)表达升高,并且已与神经原纤维缠结病理相关联。一致地,最近一项异种移植研究报告称,在人单核细胞植入人源化小鼠大脑后,同一 MG5 簇内出现了 MDM 样特征。将我们的核心 MDM 特征应用于该异种移植数据集,结果显示其在 GPNMB 巨噬细胞亚群中富集最强(图 S2O 和 S2P)。总之,这些发现表明,人类小胶质细胞的某些亚群在转录水平上与 MDM 样程序一致。
为了在蛋白水平验证 YSM 与 MDM 之间的转录差异,我们对 DR‑3x 处理的 Flt3Cre;R26‑YFP 小鼠进行了流式细胞术分析。与 YSM 相比,MDM 表达较低水平的 fractalkine 受体 CX3CR1、整合素 CD11b 和 MER 原癌基因酪氨酸激酶,以及较高水平的 CD45 和 F4/80(图 2O、2P 和 S2Q)。为了评估这些表型的稳定性,我们在植入后 4 个月通过流式细胞术分析了 DR‑3x 处理的小鼠(图 S2R)。未经处理的小鼠实质中仅含有 CD45medCD11b⁺CX3CR1⁺ 小胶质细胞,而 DR‑3x 处理的小鼠在实质中保留了一个独特的 CD45highCD11blowCX3CR1low 实质巨噬细胞群体,该群体很可能由谱系追踪实验中标记的 MDM 贡献(图 2O)。总之,这些数据表明 YSM 与 MDM 在形态、转录谱和表面标志物表达上均存在差异,并且脑内植入的 MDM 能够长期存在,尽管处于相同的组织环境中,但仍维持着独特的表面表型。
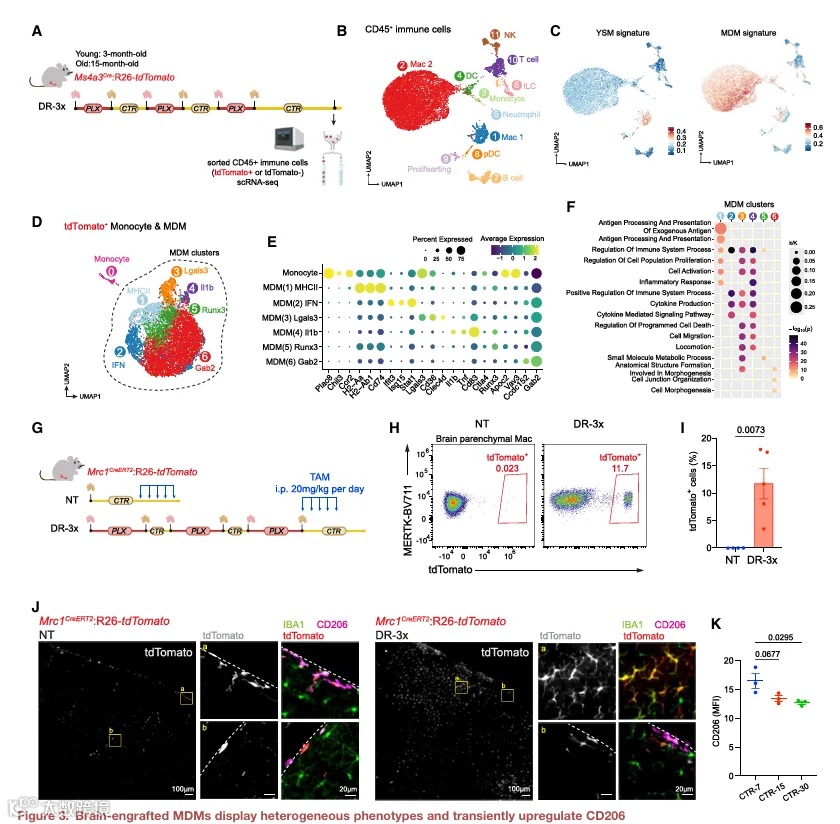
3.单细胞转录组分析揭示脑内植入 MDM 的异质性
DR‑3x 范式不仅允许 YSM 被 MDM 有效替代,而且还提供了一个无辐射模型来研究植入细胞的异质性。为了确定脑内植入的 MDM 在进入大脑后是否采用不同的转录状态,我们对从 DR‑3x 处理后的 Ms4a3Cre;R26‑tdTomato 小鼠中分离的 CD45⁺ 免疫细胞进行了 scRNA‑seq(图 3A)。为了评估 MDM 植入是否也发生在老年动物中以及年龄是否影响植入 MDM 的转录程序,我们在此分析中纳入了另外一组 15 月龄小鼠。
对所有 CD45⁺ 脑免疫细胞的无监督聚类揭示了一个多样化的免疫图谱,包括树突状细胞、T 细胞、B 细胞、固有淋巴样细胞以及两个巨噬细胞亚群 Mac1 和 Mac2(图 3B 和 S3A;表 S2)。相关性分析表明,植入老年脑的 MDM 在转录上与年轻脑中的 MDM 相似(图 S3B),表明年龄相关影响有限。模块评分分析显示,Mac1 具有最高的 YSM 评分,而 Mac2 则显示出对核心 MDM 特征的最强富集(图 3C)。这两个亚群均表达典型的小胶质细胞标志物,如 Csf1r、Tmem119 和 P2y12;然而,Sall1 的表达仅限于 Mac1,而 Mac2 则选择性地表达 BAM 基因,如 Mrc1(图 S3C)。
为了进一步解析 MDM 的异质性,我们对 tdTomato⁺ 单核细胞和 MDM 进行了亚聚类,鉴定出六个转录上不同的 MDM 亚群(图 3D‑3F)。MDM(1) 表现出高水平的抗原呈递基因(Cd74 和 H2‑Aa)。MDM(2) 富集 IFN 刺激基因(Ifit3、Isg15 和 Stat1)。MDM(3) 上调脂质处理和代谢相关基因(Lgals3 和 Cd36)。MDM(4) 显示出强烈的促炎特征,Il1b、Tnf 和 Cd83 升高,类似于在神经退行性背景下描述的单核细胞来源的疾病炎症性巨噬细胞群体。MDM(6) 主要表达与细胞连接组织和形态发生相关的基因,提示其在结构重塑中发挥作用。伪时间轨迹分析表明,MDM(1) 代表了从单核细胞到成熟 MDM 的早期过渡状态(图 S3D)。值得注意的是,MDM 亚群比例在年轻和老年脑中相似(图 S3E)。支持这一观察的是,对两种条件下 YSM 和 MDM 的 bulk RNA‑seq 分析显示,来自年轻和老年小鼠的 MDM 紧密聚集在一起(图 S3F 和 S3G),强调发育起源而非年龄是脑内植入 MDM 转录身份的主要决定因素。
我们的转录组分析显示,与 YSM 相比,MDM 中 Mrc1 的表达更高(图 2H 和 S3C)。Mrc1 编码甘露糖受体 CD206,该受体在硬脑膜巨噬细胞和 BAM 中高表达,但通常不在成体小胶质细胞中表达。有趣的是,CNS 巨噬细胞的 YS 来源前体在早期脑定植期间瞬时表达 CD206,并在进入脑实质时下调该标志物以获取小胶质细胞表型,而脑膜中的巨噬细胞后代则保留 CD206 表达。
为了检测脑内植入 MDM 是否同样瞬时表达 CD206,我们生成了 Mrc1creERT2;R26‑tdTomato 小鼠,以实现对 Mrc1 表达细胞的瞬时标记(图 3G 和 S3H)。在对照 Mrc1creERT2;R26‑tdTomato 小鼠中,他莫昔芬诱导了对硬脑膜巨噬细胞和 BAM 的有效标记,但未标记小胶质细胞或血液单核细胞(图 3H、3I 和 S3I‑S3K),tdTomato⁺ 细胞仅定位于血管周围和软脑膜区室,并与 CD206 染色共定位(图 3J)。相比之下,DR‑3x 后给予他莫昔芬导致 tdTomato⁺ 实质巨噬细胞显著增加(NT = 0.03% ± 0.01%,DR‑3x = 11.76% ± 2.77%)(图 3H 和 3I),免疫荧光证实实质内存在 tdTomato⁺ IBA1⁺ 细胞(图 3J 和 S3L)。这些细胞表现出高 CD45 和低 CX3CR1 及 CD11b 表达,与 MDM 表型一致(图 S3M)。重要的是,在分析时,实质内的 tdTomato⁺ 细胞缺乏 CD206(图 3J),表明 CD206 在植入过程中瞬时表达。DR‑3x 后的时间进程分析显示,从 CTR‑7 到 CTR‑30,CD206 信号逐渐减少(图 3K 和 S3N)。
为了测试再群体化的脑膜巨噬细胞是否直接贡献于再生的实质巨噬细胞池,我们使用 Lyve1creERT2;R26‑tdTomato 小鼠对 Lyve1⁺ 脑膜巨噬细胞进行了命运追踪(图 S3O 和 S3P)。DR‑3x 后,在脑实质中未检测到 tdTomato⁺ 细胞(图 S3Q),表明 Lyve1⁺ 脑膜巨噬细胞不浸润实质。总之,这些数据表明,脑内植入的 MDM 在进入时瞬时上调 CD206,类似于发育中的 CNS 巨噬细胞前体,而 Lyve1⁺ 脑膜巨噬细胞不供应实质巨噬细胞池。
5.YSM 与脑内植入 MDM 的独特表观遗传程序
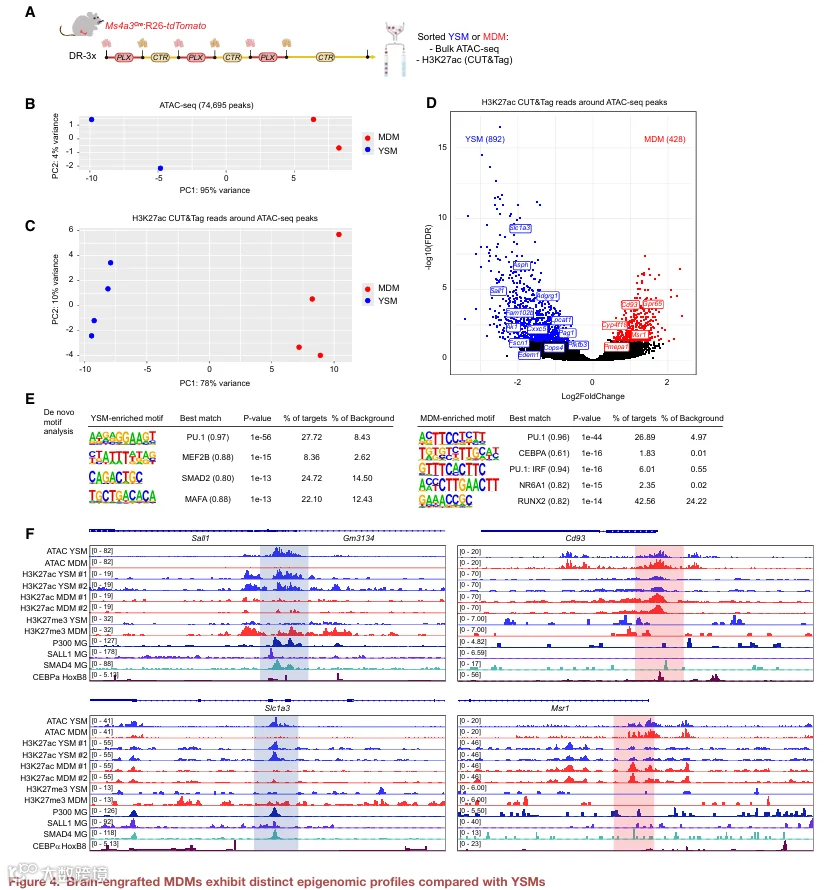
为了确定驱动 YSM 与脑内植入 MDM 不同转录状态的关键转录因子,我们分别使用 ATAC‑seq 和 H3K27ac CUT&Tag 分析了染色质可及性和组蛋白乙酰化。通过对分选的 YSM 和 MDM 中这些数据集的整合,我们能够推断与活性增强子和启动子相关的谱系特异性调控程序(图 4A)。对 74,695 个峰上的 ATAC‑seq 信号进行 PCA 分析,将 YSM 与 MDM 分开,表明两者具有不同的可及性景观(图 4B)。对 ATAC 峰周围 H3K27ac CUT&Tag 读段的 PCA 分析也区分了这两个群体,表明 YSM 与 MDM 之间的增强子/启动子激活存在差异(图 4C)。对 ATAC 定义的开放区域附近 H3K27ac 信号的差异分析确定了 892 个 YSM 特异性和 428 个 MDM 特异性乙酰化区域(图 4D)。为了推断与这些谱系偏倚调控元件相关的转录因子,我们对差异乙酰化的开放染色质峰进行了从头基序富集分析。正如预期,YSM 和 MDM 特异性峰均富集 PU.1 基序,但在其他调控输入上存在差异:YSM 特异性峰独特地富集 MEF2B、SMAD2 和 MAFA 基序,而 MDM 特异性峰则优先包含 CEBPA、PU.1:IRF 复合基序、NR6A1 和 RUNX2 基序(图 4E)。YSM 中 MEF2 和 SMAD 基序的富集值得注意,因为关键的小胶质细胞转录因子 SALL1 结合 MEF2 位点并与 SMAD 共结合以建立小胶质细胞身份。一致地,Sall1 在 YSM 中高表达,而在 MDM 中缺失(图 2I 和 S3C),并且 Sall1 启动子和超级增强子在 MDM 中不可及且以 H3K27me3 为标志(图 4F 和 S4A)。此外,一些 YSM 特异性乙酰化区域,如 Sall1 启动子、Sall1 超级增强子和 Slc1a3 启动子,确实被 SALL1 和 SMAD4 结合(图 4F 和 S4A)。相比之下,MDM 显示出更强的 C/EBP、PU.1:IRF、NR6A1 和 RUNX 模块参与,例如在 Cd93 启动子周围 H3K27ac 升高,该启动子可在 HoxB8 细胞中结合 C/EBPα(图 4F)。支持跨物种保守性的是,对人小胶质细胞亚群的转录因子预测将 RUNX2、IRF8 和 CEBPD 鉴定为 MG5 的候选上游调控因子(图 S4B),而 MG5 具有较高的 MDM 模块评分。总之,我们的数据支持一个模型,其中 SALL1‑SMAD 轴在 YSM 中维持小胶质细胞表观基因组程序,而 MDM 则采用一个以 C/EBP/IRF/RUNX 为中心的独特调控回路,该回路是其功能状态的基础。
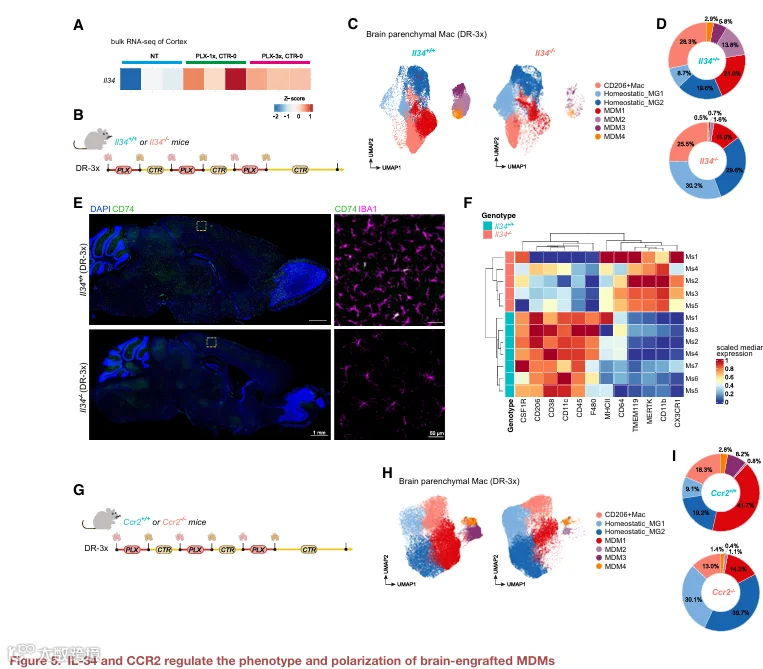
6.IL‑34 和 CCR2 调节脑内植入 MDM
接下来,我们研究了调节脑内植入 MDM 积累和表型的分子通路,重点关注微环境生存信号和招募受体。脑巨噬细胞的存活依赖 CSF1R,YSM 和 MDM 均表达 Csf1r 转录本(图 S3C)。在 CSF1R 配体中,IL‑34 对小胶质细胞发育至关重要,并由脑内的神经元和基质细胞产生。值得注意的是,PLX 处理后 IL34 表达增加(图 5A),表明 IL‑34 可能支持再群体化巨噬细胞。为了验证这一点,我们对 IL34⁺/⁺ 和 IL34⁻/⁻ 小鼠进行了 DR‑3x 处理,并通过流式细胞术分析了小鼠皮层的巨噬细胞(图 5B)。DR‑3x 后,IL34⁻/⁻ 脑内实质巨噬细胞总数减少,且未能表现出在 IL34⁺/⁺ 小鼠中观察到的 MDM 浸润特征的 CD45highCD11blow 转变(图 S4C 和 S4D)。对高维流式数据进行无监督聚类,鉴定出六个巨噬细胞簇(图 5C、S4E 和 S4F)。两个簇对应于稳态小胶质细胞(MG1 和 MG2),其特征是高表达 TMEM119 和 CD11b。一个 CD206⁺ 巨噬细胞簇表达 CD206 和 F4/80,CD45 和 TMEM119 呈中等水平,与过渡性 MDM 样状态一致。其余簇代表 MDM,其定义为高表达 CD206、F4/80 和 CD45,低表达 MERTK、CD11b、TMEM119 和 CX3CR1。其中,MDM1 为 MHC II 类低表达,MDM3 显示 CSF1R 降低且 CD11c 升高,而 MDM2 高表达 CD38(图 S4F)。在 IL34⁻/⁻ 脑中,MDM 簇(MDM1‑3)几乎完全缺失,而 CD206⁺ 巨噬细胞簇也减少,同时 MG1 和 MG2 簇增加(图 5D)。这些结果表明,IL‑34 是脑内 MDM 植入所必需的。
鉴于 CCR2 在单核细胞招募中的已知作用,我们接下来检测了 CCR2 缺失对 MDM 植入的影响(图 5E)。在 DR‑3x 处理后的 CCR2⁻/⁻ 小鼠中,MDM 簇(MDM1‑3)显著减少,但未完全消失,表明 CCR2 在促进 MDM 植入中发挥作用(图 5F、5G 和 S4G)。为了探究 CCR2 是否影响 MDM 表型,我们对 IL34⁺/⁺ 和 IL34⁻/⁻ 小鼠以及 CCR2⁺/⁺ 和 CCR2⁻/⁻ 小鼠中的 MDM 进行了差异表达分析(图 5H‑5K)。IL‑34 缺失导致 MDM 中与细胞周期、DNA 修复和代谢相关的基因下调,同时上调与抗原呈递和 IFN‑γ 应答相关的基因(图 5H 和 5I)。CCR2 缺失导致 MDM 中与炎症和趋化性相关的基因下调,同时上调与补体激活和吞噬作用相关的基因(图 5J 和 5K)。这些数据表明,IL‑34 和 CCR2 不仅调节 MDM 的丰度,还调节其表型极化。
7.血液与颅骨来源的单核细胞均可补充脑实质巨噬细胞
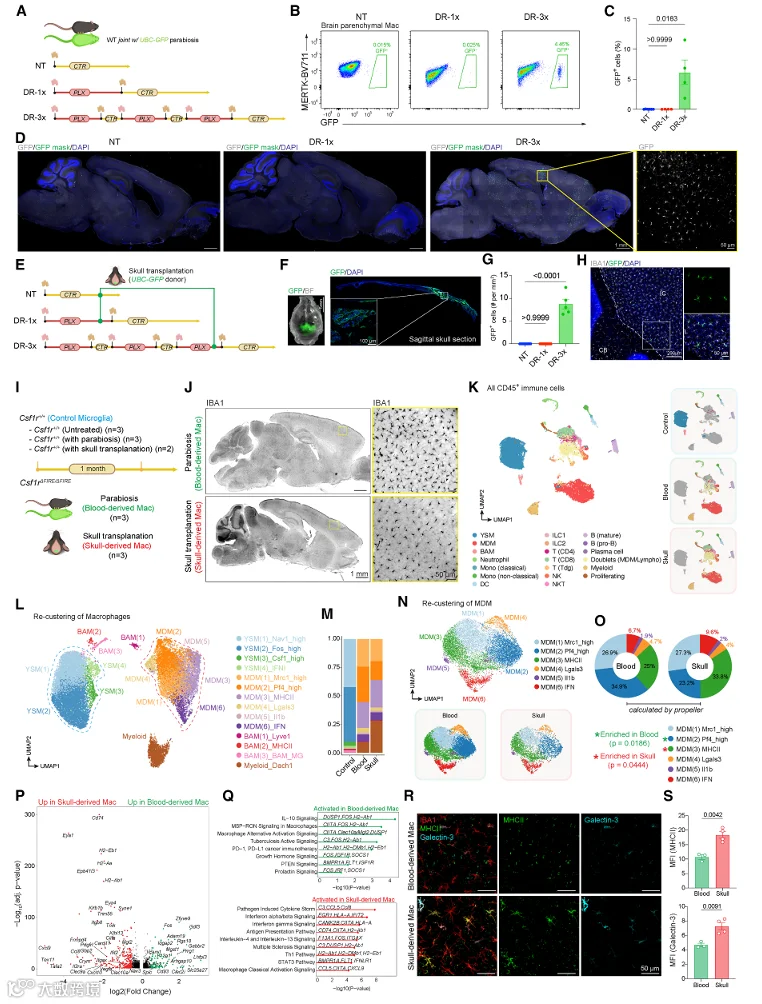
我们之前的研究表明,颅骨骨髓可以直接向硬脑膜提供免疫细胞。为了确定颅骨骨髓是否也能补充实质巨噬细胞池,我们进行了两种互补的追踪实验。首先,我们将 UBC‑GFP 小鼠与 WT 小鼠进行联体共生,以追踪循环血液来源的细胞(图 6A)。在联体共生 3 个月后,我们检测到少量 GFP⁺ 脑实质巨噬细胞,表明血液来源的单核细胞在稳态下对实质巨噬细胞池有微小贡献(图 6B 和 6C)。然而,当联体共生对随后进行 DR‑3x 处理时,GFP⁺ 脑实质巨噬细胞的百分比显著增加,表明在微环境空缺时,血液来源的单核细胞可以大量植入(图 6D‑6F)。接下来,我们通过将 UBC‑GFP 小鼠的颅骨骨瓣移植到 WT 小鼠的颅骨中,特异性地追踪颅骨来源的细胞(图 6G)。在移植后 1 个月,我们在移植骨瓣下方的脑实质中检测到 GFP⁺ 细胞,表明颅骨骨髓可以直接向脑实质输送单核细胞(图 6H 和 6I)。当受体小鼠随后进行 DR‑3x 处理时,移植骨瓣下方的 GFP⁺ 实质巨噬细胞数量显著增加(图 6J‑6L)。这些数据共同表明,血液和颅骨骨髓都是脑内 MDM 植入的潜在来源。
为了比较血液来源和颅骨来源 MDM 的分子身份,我们利用了 Csf1rΔFIRE/ΔFIRE 小鼠。该小鼠在 Csf1r 基因座中缺失 FIRE 增强子,导致组织驻留巨噬细胞(包括小胶质细胞)减少,但循环单核细胞数量不受影响。我们在 Csf1rΔFIRE/ΔFIRE 小鼠及其同窝对照中进行了 scRNA‑seq,并利用细胞哈希技术区分了血液来源和颅骨来源的 MDM(图 6M)。无监督聚类鉴定出多个巨噬细胞亚群,包括 YSM、BAM 以及两个 MDM 亚群(MDM_Blood 和 MDM_Skull)(图 6N 和 S5A)。MDM_Blood 和 MDM_Skull 均表达核心 MDM 特征基因,如 Mrc1、Cd74 和 Axl,但呈现出不同的转录状态(图 6O 和 S5B)。MDM_Skull 富集与 MHC II 类抗原呈递(H2‑Aa、H2‑Ab1)和吞噬作用(Lgals3、Cd68)相关的基因,而 MDM_Blood 则富集与细胞迁移(Ccr2、Cx3cr1)和炎症反应(Il1b、Tnf)相关的基因(图 6P 和 S5C)。这些数据表明,血液与颅骨来源的 MDM 尽管共享核心特征,但在脑实质中呈现出部分不同的转录状态。
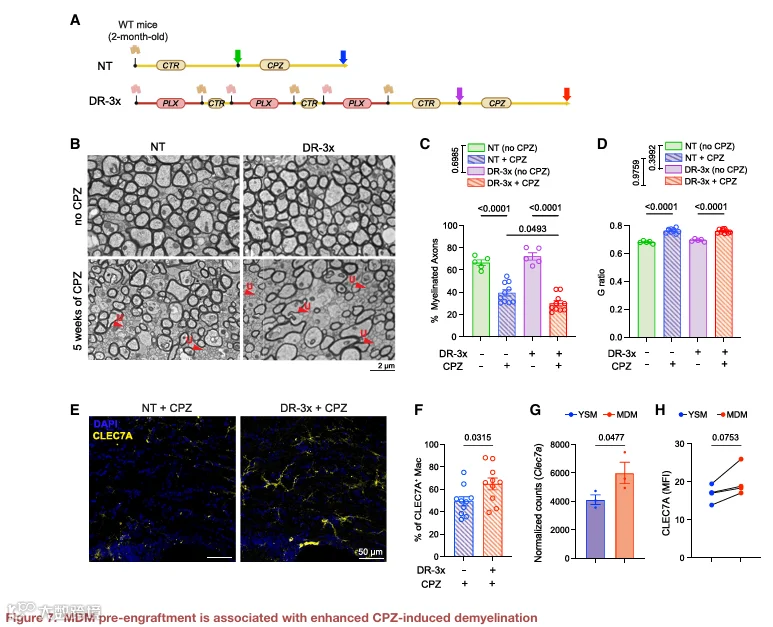
鉴于脑内植入 MDM 表现出独特的转录和表型特征,我们试图探究其存在是否对 CNS 病理具有功能性影响。我们选择了铜腙诱导的脱髓鞘模型,该模型可导致胼胝体广泛脱髓鞘。首先,我们评估了 DR‑3x 诱导的 MDM 植入对稳态髓鞘的影响。在未经处理的铜腙喂养小鼠中,胼胝体中出现广泛的髓鞘丢失和激活的 IBA1⁺ 巨噬细胞浸润(图 S6A 和 S6B)。相比之下,在铜腙饮食开始前接受 DR‑3x 处理的小鼠中,髓鞘丢失更为严重,且 IBA1⁺ 细胞浸润增加(图 S6C 和 S6D)。透射电子显微镜分析显示,与对照小鼠相比,DR‑3x 处理小鼠的胼胝体中髓鞘轴突百分比显著降低,而 G‑ratio 无显著差异(图 S6E‑S6G)。这些数据表明,MDM 植入会加剧铜腙诱导的脱髓鞘。
为了进一步探究 MDM 在脱髓鞘过程中的作用,我们利用 Ms4a3Cre;R26‑tdTomato 小鼠在 DR‑3x 后追踪 MDM,随后进行铜腙饮食(图 7A)。流式细胞术分析显示,在脱髓鞘条件下,tdTomato⁺ MDM 显著扩增,占实质巨噬细胞的比例从稳态下的约 50% 增加到铜腙处理后的 70% 以上(图 7B 和 7C)。免疫荧光分析显示,tdTomato⁺ MDM 与髓鞘碎片标志物共定位,并表达 Galectin‑3,这是一种与吞噬活性相关的蛋白(图 7D 和 7E)。为了确定 MDM 在脱髓鞘中的功能作用,我们在铜腙处理期间使用抗 CCR2 抗体阻断单核细胞招募(图 7F)。CCR2 阻断显著减少了脑内 MDM 的数量,并减轻了 DR‑3x 处理小鼠中观察到的髓鞘丢失加剧(图 7G‑7I)。总之,这些数据表明,MDM 植入会加剧铜腙诱导的脱髓鞘,而阻断 MDM 招募可减轻这种效应。
更多结果和补充图表:doi:10.1016/j.neuron.2026.01.032
长按二维码关注我们,用最短的时间和最高的效率学习更多数据分析方法!
扫描上方二维码或登录平台官网后添加CNSknowall客服微信咨询!官网地址:
https://cnsknowall.com
CNSknowall:24年最新问世的遥遥领先的科研数据(0代码生信+统计学)分析平台,同时含有机制图模块+汉化版Pubmed融合Deepseek高效筛选目标文献+SCI文献例句/语料检索模块+OPenAI官方GPT接口,>500款CNS级别图表皆可一秒内一键出图,登录即秒变数据分析大神,体验前所未有的便捷数据分析之旅,开启科研天骄之路!
可向下滑动批阅!