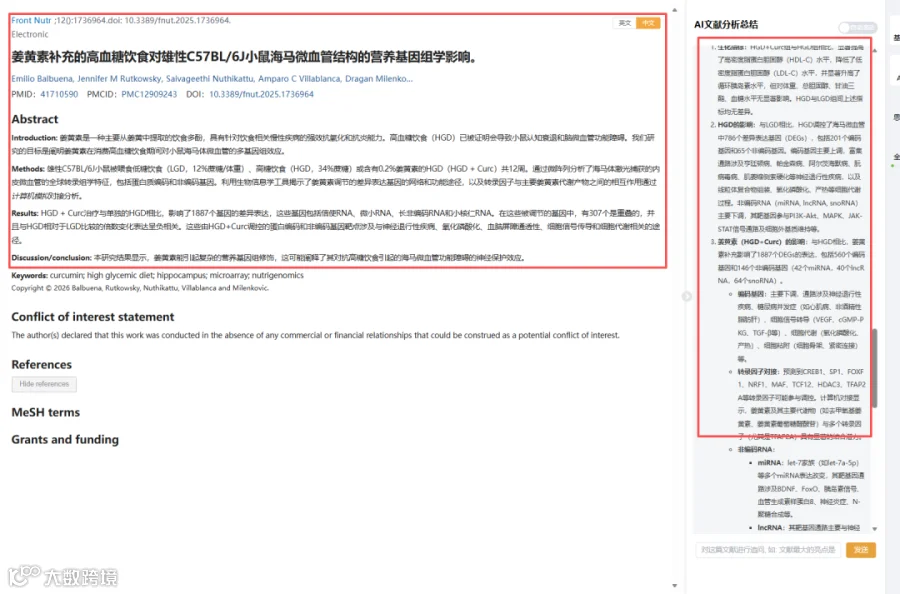
高血糖饮食(HGD)与认知功能下降密切相关,而姜黄素具有强大的抗氧化和抗炎特性。本研究首次通过激光捕获显微切割技术获取小鼠海马微血管内皮细胞,结合微阵列芯片进行多组学分析,全面解析姜黄素(0.2%膳食补充)对HGD诱导的基因表达改变的影响。研究发现,HGD+Curc组与HGD组相比,共有1887个差异表达基因,包括mRNA、miRNA、lncRNA和snoRNA。这些基因主要富集于神经退行性疾病、氧化磷酸化、血脑屏障通透性和细胞代谢等通路。特别值得注意的是,有307个被HGD上调的基因在姜黄素补充后被逆转下调。转录因子分析结合分子对接显示,姜黄素代谢物与TFAP2A、NRF1等转录因子具有显著结合能力。本研究揭示了姜黄素多层级营养基因组学调控机制,为其神经保护作用提供了新证据。
今天给大家解读一篇2月发表在《Frontiers in Nutrition》上的题目为“Nutrigenomic influence of a curcumin-supplemented high glycemic diet on hippocampal microvasculature in male C57BL/6J mice.”的文章。本研究旨在阐明姜黄素在雄性C57BL/6J小鼠摄入高血糖饮食期间,对海马微血管的多基因组影响。将小鼠分为低血糖饮食(LGD)、高血糖饮食(HGD)和HGD加0.2%姜黄素(HGD+Curc)三组,干预12周。通过激光捕获显微切割技术分离海马微血管,利用微阵列分析其全局转录组谱(包括编码和非编码基因)。运用生物信息学工具揭示姜黄素调控的差异表达基因所涉及的网络和功能通路,并通过计算机对接分析预测转录因子与主要姜黄素代谢物之间的相互作用。(请持续关注我们,每天为您解读最新见刊的文献!)想薅生信资料羊毛?直接在对话框回复 “资料”,免费领取干货大礼包!
不想做实验,没数据,还想要快速发表文章,没问题的!公共数据库就是我们的数据宝藏!没思路不用担心,作为专业的生信团队,我们很乐意为你们效劳,提供研究路线设计和数据挖掘分析,扫码联系我们吧!


团队成员合影(位于上海陆家嘴中心,可随时预约参观)
题目:《姜黄素补充的高血糖饮食对雄性C57BL/6J小鼠海马微血管结构的营养基因组学影响》Nutrigenomic influence of a curcumin-supplemented high glycemic diet on hippocampal microvasculature in male C57BL/6J mice
发表期刊:Frontiers in Nutrition
影响因子:5.1
研究背景:
- 问题
高血糖饮食(HGD)或西方饮食可导致认知功能下降和小鼠大脑微血管功能障碍,是代谢综合征、2型糖尿病及相关神经病变的风险因素。
- 干预物
姜黄素是姜黄中的主要多酚类物质,具有抗氧化和抗炎能力,在缓解代谢综合征相关认知障碍方面显示出潜力,但其对高血糖饮食背景下海马微血管的基因组水平影响尚不清楚。
- 研究缺口
尽管有研究关注姜黄素对整体海马或认知功能的影响,但对其在海马微血管内皮细胞中,特别是在高血糖挑战下的营养基因组学效应,在野生型模型中尚未被充分探索。
CNSknowall 平台 Pubmed+AI 快速提炼全文要点
研究思路:
- 动物实验
19周龄雄性C57BL/6J小鼠随机分为LGD、HGD、HGD+Curc三组,饲喂12周。
- 样本采集
干预结束后,采集体重、空腹血清(测血脂、血糖、胰岛素),并快速取脑。
- 靶向取样
对海马组织进行冰冻切片,通过碱性磷酸酶染色和激光捕获显微切割(LCM)特异性分离海马微血管(<20 μm,主要代表CA1和CA3区)。
- 基因组学分析
提取LCM获取的微血管总RNA,使用Clariom D Mouse微阵列进行转录组分析,覆盖蛋白编码和非编码基因。
- 生物信息学分析
对差异表达基因(DEGs)进行RNA分类、通路富集分析、转录因子预测、计算机模拟对接(姜黄素代谢物与转录因子)、非编码RNA靶基因分析、网络构建及人类疾病关联分析。
研究亮点:
- 多层次基因组分析
不仅分析了蛋白编码mRNA,还全面考察了miRNA、lncRNA、snoRNA等非编码RNA的表达变化,揭示了姜黄素多层次的基因组调控作用。
- 反向调控效应
发现姜黄素补充能显著逆转(负相关)HGD引起的307个基因的表达变化,直观展示了其对抗HGD不良影响的分子证据。
- 机制关联深入
通过生物信息学分析,将差异表达基因与神经退行性疾病、糖尿病并发症等具体病理通路直接关联,并预测了姜黄素代谢物与关键转录因子的相互作用。
- 聚焦特定脑区微血管
研究针对海马区的激光捕获微血管内皮细胞进行分析,使发现更精确地反映大脑关键记忆区域微血管的分子变化。
研究结果:
- 生化指标
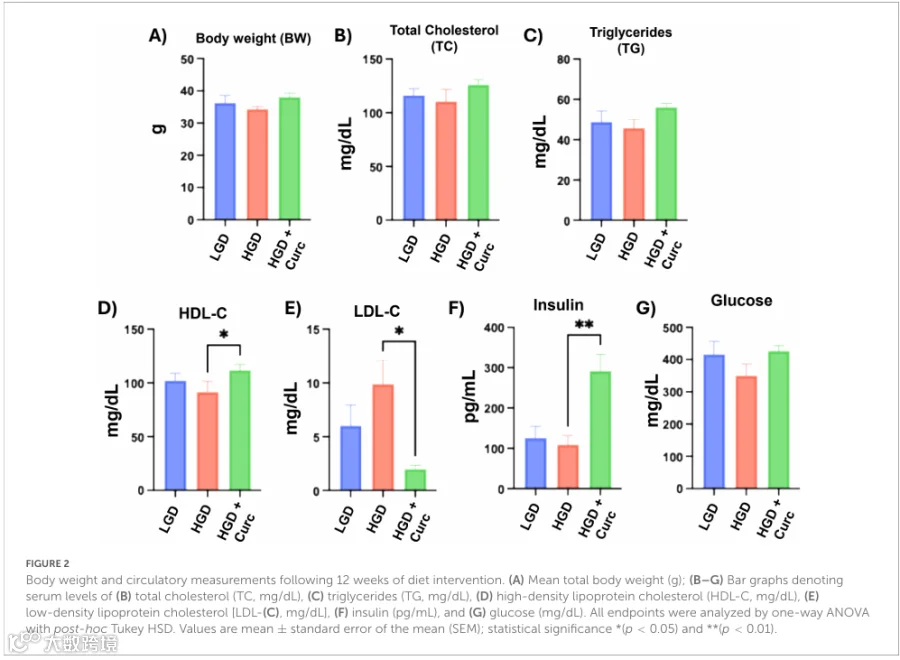
HGD+Curc组与HGD组相比,显著提高了高密度脂蛋白胆固醇(HDL-C)水平,降低了低密度脂蛋白胆固醇(LDL-C)水平,并显著升高了循环胰岛素水平,但对体重、总胆固醇、甘油三酯、血糖水平无显著影响。HGD与LGD组间上述指标均无差异。
- HGD的影响
与LGD相比,HGD调控了海马微血管中786个差异表达基因(DEGs),包括201个编码基因和65个非编码基因。编码基因主要上调,富集通路涉及亨廷顿病、帕金森病、阿尔茨海默病、朊病毒病、肌萎缩侧索硬化等神经退行性疾病,以及线粒体复合物组装、氧化磷酸化、产热等细胞代谢过程。非编码RNA(miRNA, lncRNA, snoRNA)主要下调,其靶基因参与PI3K-Akt、MAPK、JAK-STAT信号通路及细胞外基质维持等。
- 姜黄素(HGD+Curc)的影响
与HGD相比,姜黄素补充影响了1887个DEGs的表达,包括560个编码基因和146个非编码基因(42个miRNA,40个lncRNA,64个snoRNA)。
- miRNA
let-7家族(如let-7a-5p)等多个miRNA表达改变,其靶基因通路涉及BDNF、FoxO、胰岛素信号、血管生成素样蛋白8、神经炎症、N-聚糖合成等。
- lncRNA
其靶基因通路主要与神经发育障碍(如脆性X综合征、自闭症)、神经元分化、谷氨酸能突触调节、NO/cGMP/PKG神经保护等相关。
- snoRNA
部分C/D盒snoRNA(如Snord16a, Snord59a)表达发生改变。
- 编码基因
主要下调,通路涉及神经退行性疾病、糖尿病并发症(如心肌病、非酒精性脂肪肝)、细胞信号转导(VEGF、cGMP-PKG、TGF-β等)、细胞代谢(氧化磷酸化、产热)、细胞粘附(细胞骨架、紧密连接)等。
- 转录因子对接
预测到CREB1、SP1、FOXF1、NRF1、MAF、TCF12、HDAC3、TFAP2A等转录因子可能参与调控。计算机对接显示,姜黄素及其主要代谢物(如去甲氧基姜黄素、姜黄素葡萄糖醛酸苷)与多个转录因子(尤其是TFAP2A)具有显著的结合潜力。
- 非编码RNA
- 姜黄素的逆转效应
在HGD/LGD和HGD+Curc/HGD两个关键比较中,存在376个共同DEGs。其中307个基因的表达趋势在HGD+Curc组中被逆转(与HGD效应呈负相关)。这些基因同样富集于神经退行性疾病、糖尿病并发症和细胞代谢通路。这表明姜黄素能对抗HGD诱导的部分有害基因组变化。
- 疾病关联
HGD+Curc调控的DEGs在基因组学上与人类神经系统疾病、神经退行性疾病和神经发育疾病显著相关。
研究总结:
- 主要结论
本研究表明,膳食姜黄素能诱导复杂的营养基因组学修饰,通过多层面(编码mRNA、非编码RNA、潜在转录因子调控)调控海马微血管的基因表达。这些变化可能阐明其对抗高血糖饮食诱导的海马微血管功能障碍的神经保护作用。姜黄素逆转了HGD诱导的多个与神经退行性病变、线粒体功能障碍和内皮重塑相关通路的基因表达。
- 作用机制探讨
- 调控线粒体功能
下调氧化磷酸化相关基因,可能减少活性氧(ROS)的过度产生。
- 影响神经退行相关蛋白
下调淀粉样前体蛋白(App)及其相关分子、GSK3α等,可能抑制Aβ聚集和tau蛋白磷酸化。
- 保护血脑屏障
下调VEGF/PI3K/RhoA通路相关基因及基质金属蛋白酶(MMPs),可能抑制病理性血管生成和降低BBB通透性。
- 表观遗传调控
通过改变miRNA、lncRNA、snoRNA的表达,影响下游靶基因网络。
- 潜在直接作用
姜黄素代谢物可能与CREB1、TFAP2A、NRF1、HDAC3等关键转录因子结合,影响其活性。
- 潜在双重角色
研究结果提示,在本模型背景下,姜黄素可能同时具有中度神经保护和潜在抗肿瘤(通过抑制血管生成和细胞能量代谢)的双重作用。
- 局限性
研究依赖特定生物信息学工具和数据库;大量DEGs未被鉴定或归类;HGD在12周内未引起系统性高血糖或胰岛素抵抗(可能与饮食设计、干预时长有关);仅使用雄性小鼠;未直接测量Aβ等病理蛋白聚集。
- 总结论
姜黄素对海马微血管具有多层次的营养基因组学效应,主要调控神经退行性病变、线粒体功能障碍和内皮重塑等关键信号通路之间的相互作用,以应对高血糖饮食的挑战。
结果译文:
在12周饮食干预期结束时,LGD和HGD对照组之间的总体体重没有显著差异(图2A)。同样,与LGD相比,HGD对总胆固醇、甘油三酯、HDL-C、LDL-C、胰岛素和葡萄糖的血清水平没有显著影响(图2B-G)。值得注意的是,与HGD相比,补充姜黄素(HGD+Curc)显著提高了HDL-C(p < 0.05)并降低了LDL-C(p < 0.05),同时显著提高了胰岛素水平(p < 0.01)(图2D-F)。其他生化参数在HGD+Curc组与HGD组之间没有显著差异。
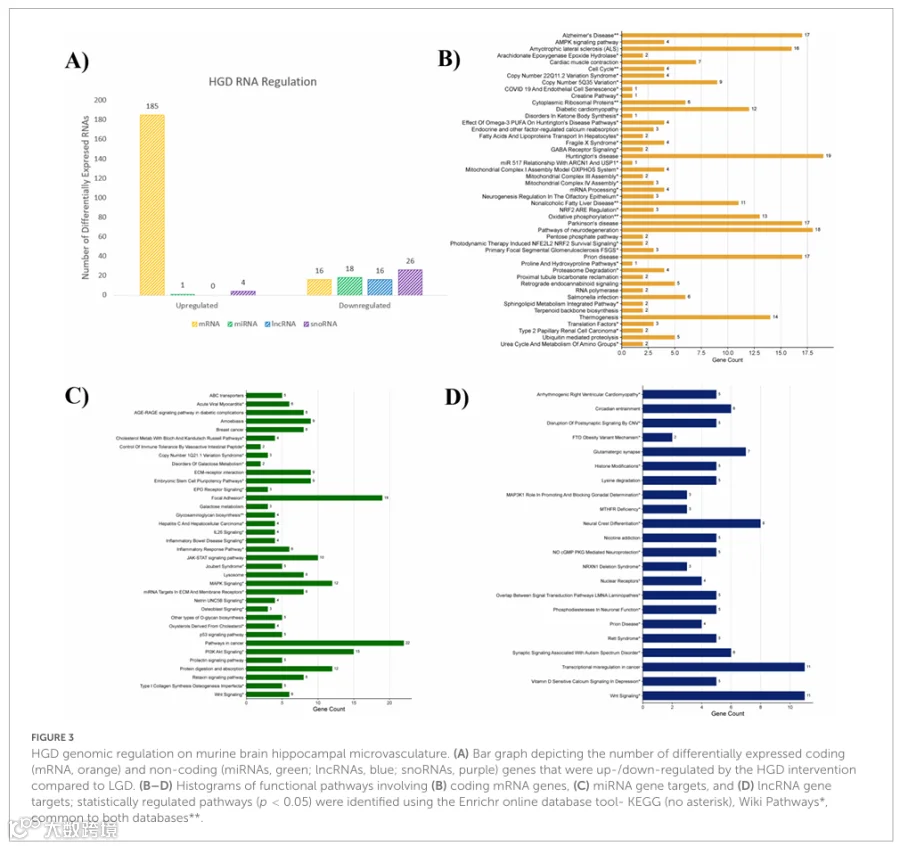
2.HGD/LGD:高血糖饮食对雄性小鼠海马微血管的影响
对微阵列数据的统计分析显示,与LGD相比,HGD处理调节了雄性小鼠海马微血管中786个差异表达基因(补充表1)。更具体地说,HGD处理调节了201个蛋白编码基因和65个非编码基因;后一类包括19个microRNA、16个长链非编码RNA和30个小核仁RNA。在这些特征基因中,与LGD相比,HGD干预上调了190个基因,下调了76个基因(图3A)。此外,剩余的520个DEG被归类为假基因、多复合体基因或未分配基因(有符号或无符号)。
在这些分类中,共有201个蛋白编码基因被HGD相对于LGD差异表达,其中大多数上调(n=185,变化倍数范围1.5-11.4),少数下调(n=16,变化倍数范围-4.38至-1.5,补充表2)。揭示基因和通路本体论的生物信息学分析表明,编码DEGs参与了主要神经退行性疾病如亨廷顿病、帕金森病、阿尔茨海默病、朊病毒病和肌萎缩侧索硬化症的通路调节,以及细胞代谢(例如线粒体复合物组装、氧化磷酸化和产热)(图3B)。
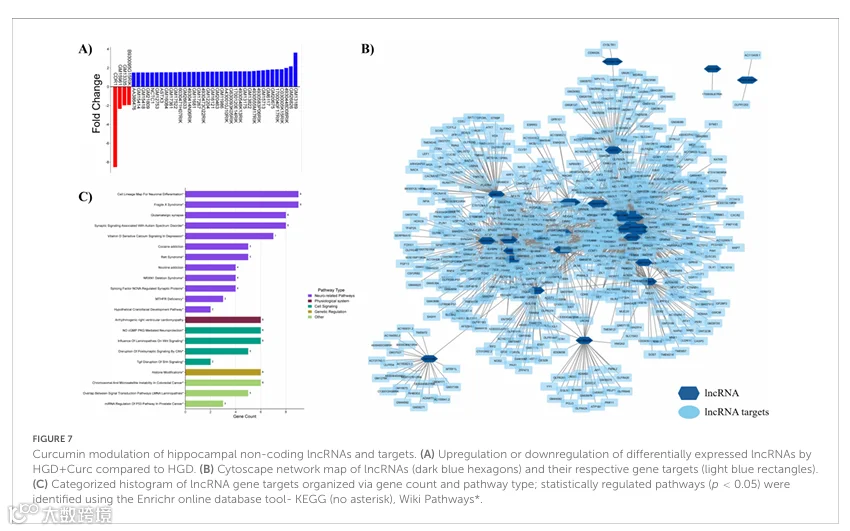
海马微阵列分析表明,与LGD相比,HGD处理也差异表达了非编码RNA,如miRNAs、lncRNAs和snoRNAs。首先,19个受调控的miRNAs主要下调(n=18,变化倍数范围-3.93至-1.5),只有一个(mmu-miR-692)上调,变化倍数为4.97(补充表3)。受调控miRNAs的靶基因共527个,参与PI3K/Akt、MAPK和JAK/STAT信号通路以及细胞外基质维持,如ECM-受体相互作用和细胞骨架黏着斑(图3C)。此外,与LGD相比,HGD下调了所有16个差异表达的lncRNAs,变化倍数范围为-6.23至-1.51(补充表4)。鉴定出的受差异表达lncRNAs调控的542个靶基因涉及神经元功能通路,如神经嵴分化、NO/cGMP/PKG介导的神经保护作用和磷酸二酯酶,以及突触信号传导(Wnt和谷氨酸能突触调节)(图3D)。HGD干预相对于LGD显著调控的所有通路(p<0.05),按KEGG和Wiki Pathways数据库分类,已提供于补充图1(编码基因,图1A;miRNA靶基因,图1B;lncRNA靶基因,图1C)。最后,30个snoRNAs的相对表达主要下调(n=26;变化倍数-3.27至-1.51),部分上调(n=4;变化倍数1.92-19.32)(补充表5)。通过文献检索和生物信息学工具未揭示这些snoRNAs的相关靶基因和通路。
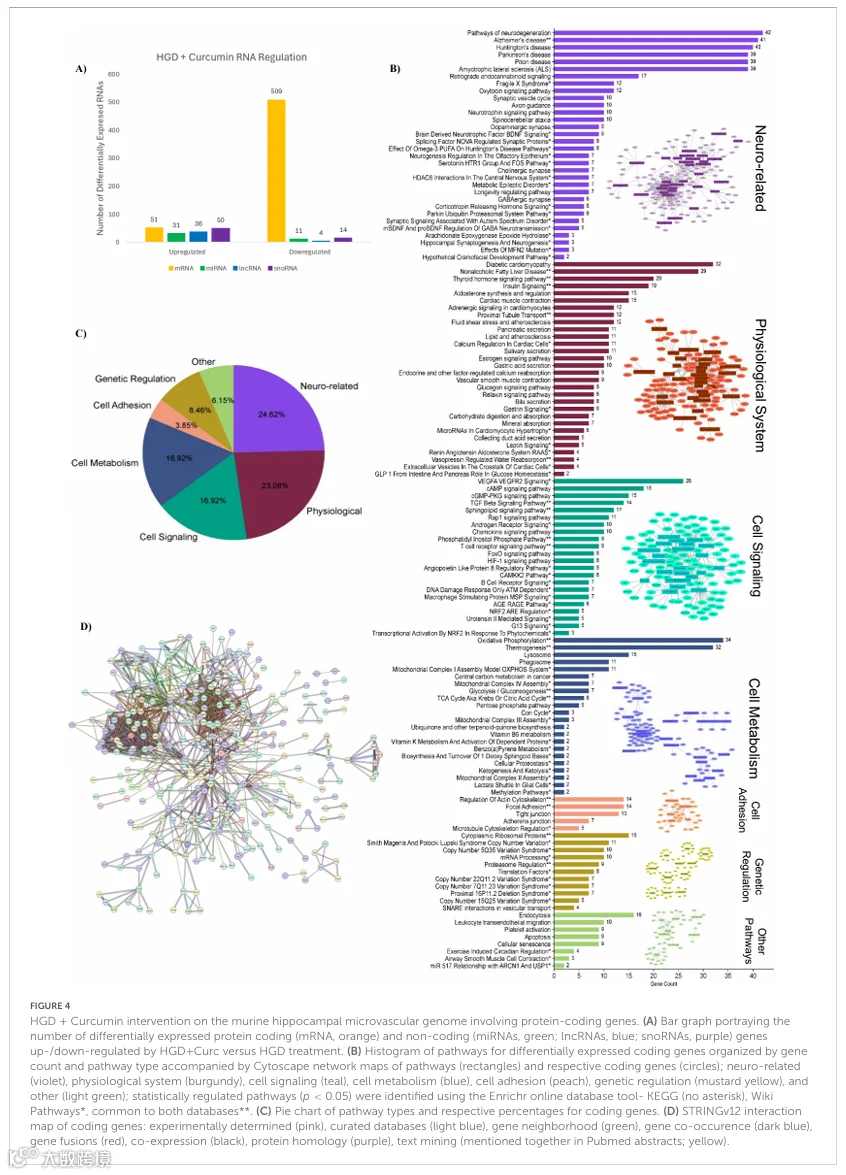
3.HGD+Curc/HGD:高血糖饮食+姜黄素对海马微血管基因组的营养保健作用
还对姜黄素补充组进行了微阵列分析,以揭示这种膳食多酚如何影响暴露于高血糖负荷的小鼠海马微血管的基因组。对微阵列数据的统计分析表明,与HGD对照组相比,HGD+Curc处理影响了1887个DEGs(补充表6)。用基因符号鉴定的DEGs包括560个蛋白编码基因和146个非编码基因(42个miRNAs,40个lncRNAs和64个snoRNAs)。在归入这些类别的基因中,与HGD相比,HGD+Curc上调了168个基因,下调了538个基因(图4A)。剩余的1181个DEGs被归类为假基因、多复合体基因、免疫球蛋白可变链基因或未分配基因(有基因符号或无符号)。
3.1 HGD+Curc对编码mRNA的影响
与HGD组相比,HGD+Curc差异表达的蛋白编码基因主要下调(n=509,变化倍数-32.64至-1.5),部分上调(n=51,变化倍数1.51-9.68)(补充表7)。通路富集分析表明,受HGD+Curc影响的编码DEGs参与了各种神经和生理系统相关功能,以及细胞信号传导、代谢、遗传调控和黏附通路(图4B)。涉及编码DEGs的认知和神经相关功能占整体显著调控通路的24.62%(图4C),主要与神经退行性疾病(如亨廷顿病、帕金森病和阿尔茨海默病;朊病毒病;ALS)以及激素(催产素、促肾上腺皮质激素)和/或生长因子如BDNF的信号传导有关。涉及其他生理过程(23.08%)的通路也被识别,如糖尿病心肌病、非酒精性脂肪性肝病以及胰岛素和甲状腺激素的信号传导。细胞转导通路(16.92%)涉及VEGF/VEGFR2、cAMP/cGMP-PKG和TGF-β信号传导,而编码DEGs在细胞代谢(16.92%)中的功能与氧化磷酸化和产热的调节密切相关。虽然占比很小但对内皮维持重要,细胞黏附相关通路(3.85%)包括肌动蛋白/微管细胞骨架的维持、黏着斑和紧密/黏附连接。遗传调控通路(8.46%)如mRNA加工、翻译因子和核糖体蛋白的调节也受到影响,这是蛋白质编码水平的营养基因组学调控的特征。此外,一些鉴定出的基因与染色体异常有关,如拷贝数变异和近端缺失,受膳食干预影响。最后,HGD+Curc调控的编码DEGs还参与了其他调控通路(6.15%),涉及细胞过程如内吞、凋亡和衰老。
STRING图谱突出了差异表达的蛋白编码基因之间的潜在相互作用(图4D)。560个编码基因中有28个参与了25次或更多相互作用(表1),其中相互作用次数最多(38次)的是ATP合酶亚基α。这些高度相互作用的编码基因参与了神经退行性疾病、糖尿病并发症(即NAFLD和心肌病)以及氧化磷酸化和产热的细胞代谢过程通路。
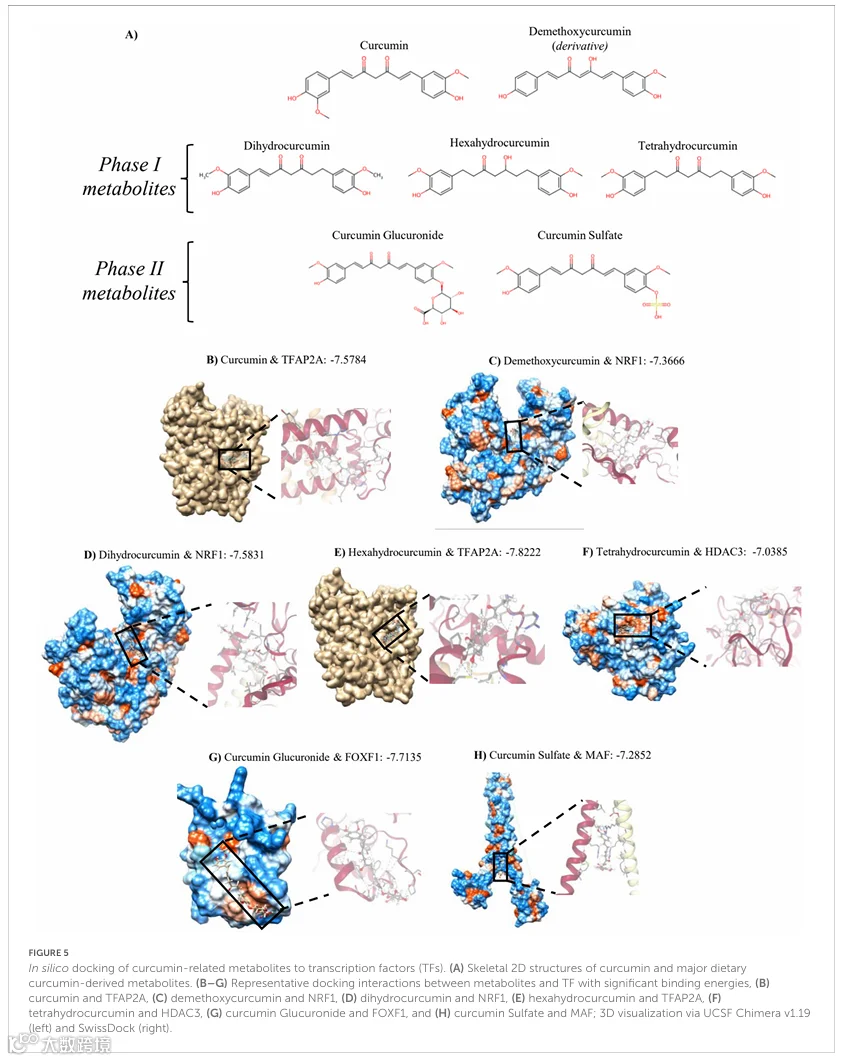
3.2 转录因子与姜黄素代谢物的计算机模拟对接
本研究中观察到的姜黄素消费的另一个营养基因组学调控水平是通过HGD+Curc调控的蛋白编码DEGs与潜在转录因子之间的相互作用,这些转录因子的活性可能受到姜黄素的影响并导致观察到的基因组变化。鉴定出七个统计学上显著的TFs(p<0.05),包括CREB1、SP1、FOXF1、NRF1、MAF、TCF12、HDAC3和TFAP2A(表2)。与CREB1相关的编码DEGs数量最多(n=33),而最显著的关系(p=0.0011)是与MAF,这可能是因为Maf本身是一个DEG。为了进一步研究姜黄素消费如何潜在地影响TF活性,进行了计算机模拟对接分析,以确定鉴定出的TFs与姜黄素的主要膳食代谢物或衍生物之间的结合能。姜黄素和相关化合物去甲氧基姜黄素、二氢姜黄素、六氢姜黄素、四氢姜黄素、姜黄素葡糖苷酸和姜黄素硫酸酯的结构如图5A所示,它们与假定的TFs的结合能见表3。大多数代谢物与一个或多个TF有显著对接(<-7 kcal/mol),最低结合能始终与TFAP2A相关,除了四氢姜黄素和NRF1。在代谢物中,去甲氧基姜黄素和姜黄素葡糖苷酸显示出与最多数量蛋白质的潜在相互作用。膳食化合物与TFs之间显著对接的代表性描绘如图5B-H所示。
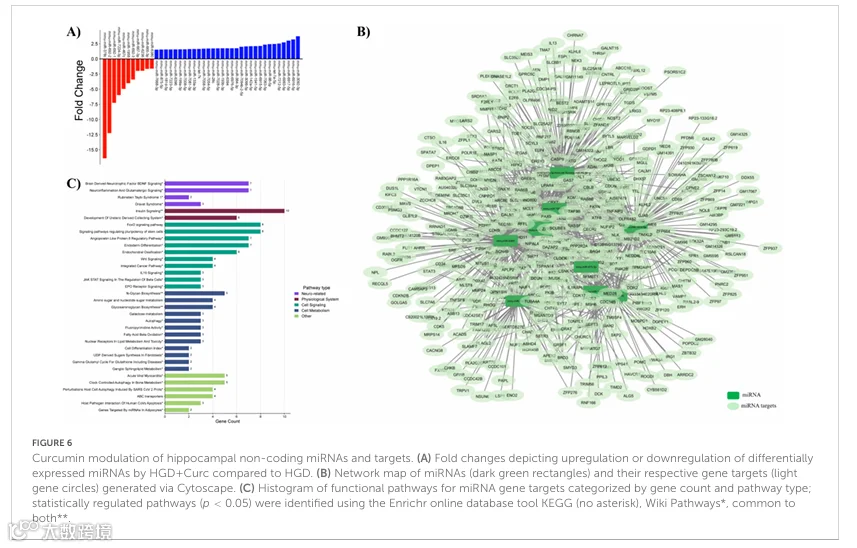
3.3 HGD+Curc非编码miRNA
与HGD一样,姜黄素补充使海马微血管中的几类非编码RNA差异表达。首先,与HGD相比,HGD+Curc调控了42个miRNAs,其中31个上调(变化倍数范围1.53-3.75),11个下调(变化倍数范围-16.41至-1.6)(图6A和补充表8A)。生物信息学分析鉴定出仅12个DE miRNAs的470个靶基因(p<0.05),其中10个miRNAs上调(变化倍数1.55-2.46),两个下调(变化倍数-16.41至-1.65)(补充图4A)。参与miRNAs和各自基因靶标的网络图如图6B所示,表明基因与多达八个miRNAs相关。特别地,三个miRNAs(mmu-let-7a-5p、let-7k和miR-98-5p)在网络节点中组合在一起,因为它们是let-7家族成员,被上调(变化倍数分别为2.46、1.65、2.44),并与最大子集(n=179)的靶基因相关(补充表8B)。DE miRNAs靶向的基因的整体功能与BDNF、FoxO和胰岛素信号传导以及血管生成素样蛋白-8调节通路有关(图6C),这些通路与前述HGD+Curc差异表达的编码基因共有。miRNA靶标独有的其他通路与干细胞多能性调节、神经炎症和谷氨酸能信号传导以及N-聚糖和糖胺聚糖的生物合成和半乳糖及氨基/核苷酸糖的代谢有关。
3.4 HGD+Curc非编码lncRNA
姜黄素补充还调控了海马微血管中的40个lncRNAs,其中大多数上调(n=36,变化倍数1.5-3.62),仅四个下调(变化倍数:-8.53至-1.93)(图7A和补充表9)。发现大多数DE lncRNAs(82.5%)共有656个基因靶标,并在图7B提供的网络图中描绘。通路富集分析显示,这些lncRNA靶标主要涉及神经功能。然而,这些受lncRNAs调控的基因与神经发育障碍相关,而非神经退行性疾病,如脆性X综合征、自闭症、雷特综合征、亚甲基四氢叶酸还原酶缺乏症、假设的颅面发育通路、拷贝数变异导致的突触后信号破坏以及T-box G-rich相互作用因子对Shh信号的破坏(图7C)。lncRNA靶基因的其他显著调控通路包括神经元分化、谷氨酸能突触调节和NO/cGMP/PKG神经保护作用。按数据库(即KEGG和Wiki Pathways)和字母顺序另行组织的、HGD+Curc与单独HGD相比显著调控的通路(p<0.05)已在补充图2-4中为编码DEGs以及miRNAs和lncRNAs的基因靶标提供。
3.5 HGD+Curc非编码snoRNA
最后,HGD+Curc干预也差异调控了非编码snoRNAs(n=64),其中50个上调(变化倍数1.5-4.72),其他14个下调(变化倍数-16.58至-1.54)(补充表10)。通过文献检索和生物信息学工具未观察到这些snoRNAs的靶基因和通路,尽管鉴定出几个C/D盒snoRNAs。值得注意的是,Snord16a和Snord59a分别被HGD+Curc下调(变化倍数:-4.07)和上调(变化倍数:1.59)。
3.6 HGD+Curc:编码DEGs与非编码基因靶标的整合分析
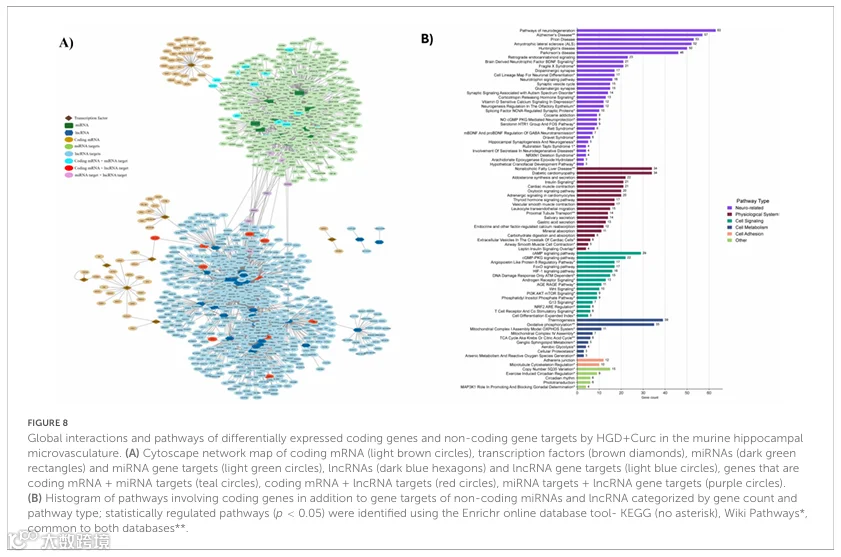
为了进一步研究HGD+Curc膳食干预对海马微血管的多基因组学影响,我们生成了一个整合网络,包括差异表达的蛋白编码mRNAs、潜在调控TFs以及非编码miRNAs和lncRNAs,以及它们各自的基因靶标(图8A)。靶标围绕TFs和非编码RNA聚集,有一些重叠,表明某些基因通过不止一种RNA类型经历多水平调控。例如,各种基因被鉴定为蛋白编码DEGs,同时也是miRNAs(n=13:Aplp2、Dram2、Epm2aip1、Erh、Ghitm、Gm14325、Gm14326、Gm14391、Nlk、Ppp2r2a、Rab6b、Slc25a18、Vampl)或lncRNAs(n=10:Agap2、Arf3、Atp1b1、Atp2b4、Camk2n1、Cask、Epb4111、Lgi1、Srp54a、Tgfb3)的靶标(补充图5A)。同样,少数基因是miRNAs和lncRNAs的共同靶标(n=8:Baz2a、Brwd3、Caln1、Ccnt1、Dnal1、Slitrk2、Trim56、Yy1),尽管通过生物信息学工具未鉴定出任何基因在编码DEGs和非编码多调控靶标(即所有三组)中共同存在。
对所有差异表达的编码基因以及miRNAs和lncRNAs的基因靶标进行了整合通路富集分析,以深入了解HGD+Curc干预对海马微血管诱导的整体、多基因组调控(图8B)。一致地,编码DEGs和非编码基因靶标的很大一部分参与了神经退行性疾病通路以及BDNF和逆行内源性大麻素神经元信号传导。此外,糖尿病后遗症如心肌病、NAFLD以及胰岛素和醛固酮的相互作用受到HGD+Curc消费的影响。几种转导通路如cAMP/cGMP-PKG、FoxO、HIF-1和血管生成素样蛋白8信号传导以及产热和氧化磷酸化的细胞代谢过程被显著调控。至于内皮黏附维持,涉及微管和黏附连接调节的基因被识别。最后,一些DEGs参与了调控拷贝数变异、昼夜节律和光转导的其他通路。HGD和HGD+Curc之间通路的重叠以及共同的编码DEGs和/或非编码基因靶标分别在补充图5B和C中提供。
3.7 HGD+Curc差异表达基因与疾病的关联
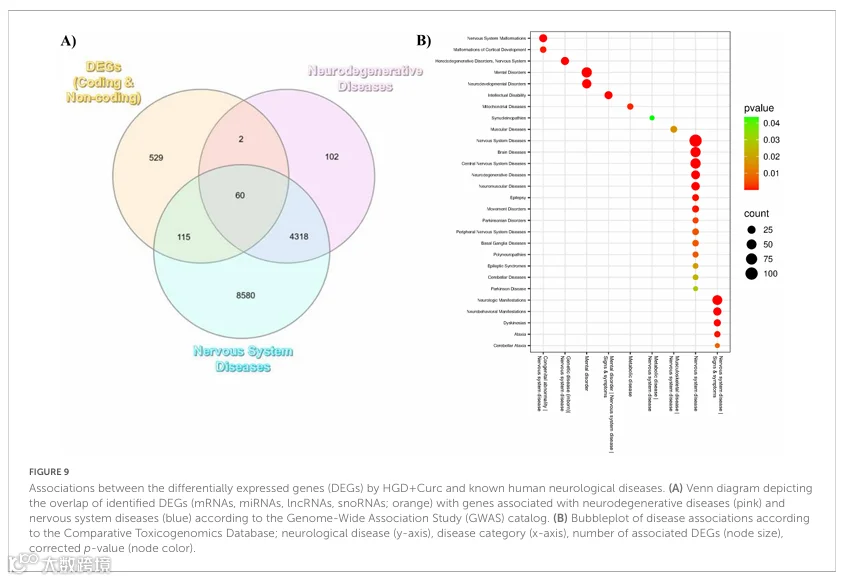
由于通过通路富集分析发现鉴定的DEGs持续参与神经功能障碍,我们对小鼠海马基因组中鉴定的差异表达基因与已知参与人类神经系统疾病发展的基因之间的基因组关联进行了进一步研究。根据GWAS目录,HGD+Curc调控的编码和非编码DEGs与人类神经系统疾病(n=115)、神经退行性疾病(n=2)或两者兼有(n=60)中观察到的调控基因重叠(图9A)。通过CTD更仔细地观察,HGD+Curc调控的DEGs与一般神经系统疾病高度相关(校正后p=6.28×10⁻²⁴),以及几个亚类如先天性异常、精神障碍和遗传性、代谢性和肌肉骨骼疾病(图9B和补充表11)。值得注意的是,HGD+Curc相关的DEGs进一步显示与神经退行性疾病(校正后p=2.48×10⁻⁹)和神经发育障碍(校正后p=5.02×10⁻⁹)相关。还观察到与神经系统疾病体征和症状的关联,如智力障碍、神经/神经行为表现、运动障碍和共济失调。
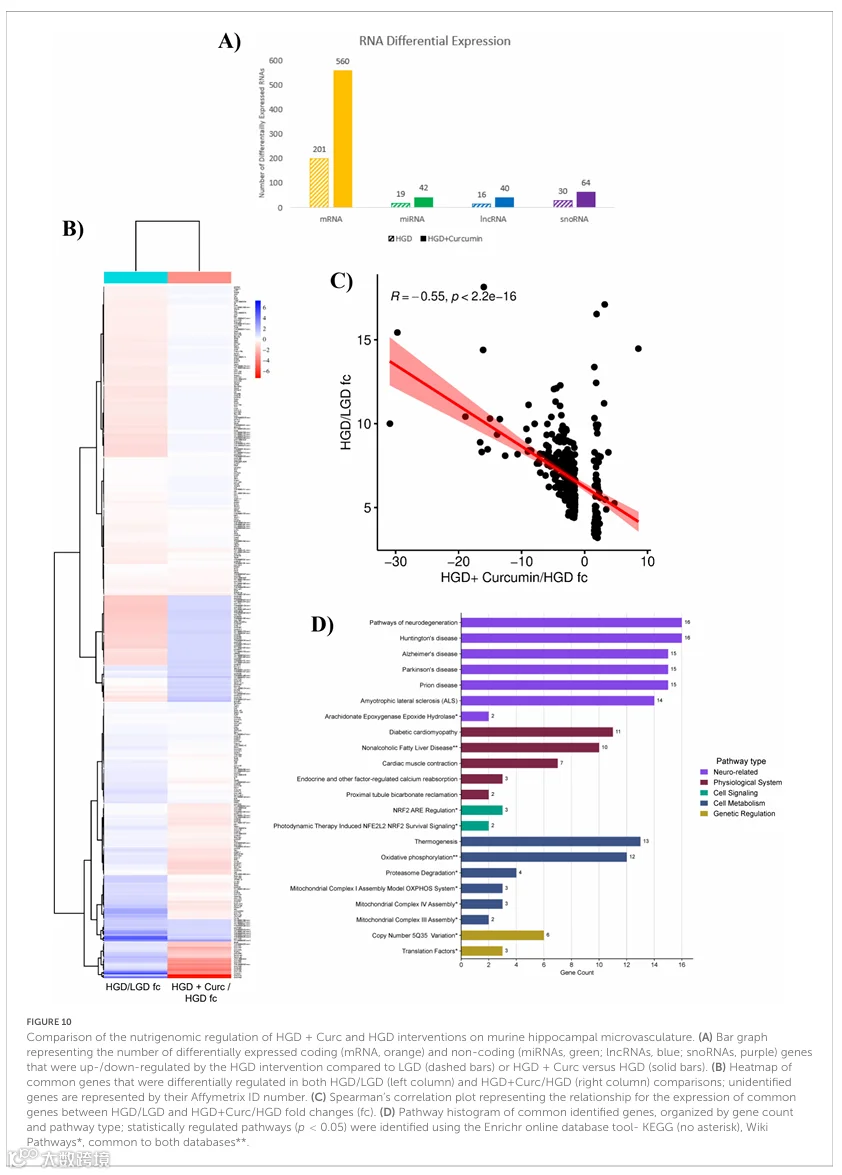
3.8 姜黄素抵消HGD诱导的差异基因表达
本研究的总体目标是确定膳食干预组之间小鼠脑微血管的差异基因组调控,其中两个关键比较是HGD与LGD以及HGD+Curc与HGD。如图10A所示,相对于HGD,HGD+Curc补充组差异表达的蛋白编码基因(560 vs. 201)和非编码基因(146 vs. 65)数量是HGD和LGD两个对照组之间鉴定出的DEGs的两倍以上。两个关键比较中共有376个共同DEGs,其中包括139个编码mRNAs、5个miRNAs、4个lncRNAs和14个snoRNAs,其余是杂项或未鉴定基因(图10B和补充表12)。所有这些共同DEGs在HGD相对于LGD中具有上调的变化倍数(3.2-18.14),而其中307个在HGD+Curc相对于HGD中表达逆转下调(-30.91至-1.5),剩下69个上调(1.5至8.53)。此外,相关性分析计算了两个关键比较HGD/LGD和HGD+Curc/HGD之间共同DEGs变化倍数的显著负相关(p < 2.2×10⁻¹⁶;R = -0.55)(图10C),提示姜黄素的抵消作用。通路富集分析(图10D)表明这些共同DEGs参与了神经退行性疾病、糖尿病并发症以及细胞代谢过程(如线粒体复合物组装、氧化磷酸化和产热)。
更多结果和补充图表:doi:10.3389/fnut.2025.1736964
长按二维码关注我们,用最短的时间和最高的效率学习更多数据分析方法!
扫描上方二维码或登录平台官网后添加CNSknowall客服微信咨询!官网地址:
https://cnsknowall.com
CNSknowall:24年最新问世的遥遥领先的科研数据(0代码生信+统计学)分析平台,同时含有机制图模块+汉化版Pubmed融合Deepseek高效筛选目标文献+SCI文献例句/语料检索模块+OPenAI官方GPT接口,>500款CNS级别图表皆可一秒内一键出图,登录即秒变数据分析大神,体验前所未有的便捷数据分析之旅,开启科研天骄之路!
可向下滑动批阅!