本研究通过单细胞RNA测序及空间转录组学分析新辅助免疫治疗胃癌患者样本,鉴定出肿瘤内源性YAP1是驱动免疫治疗抵抗的关键因子。YAP1通过上调CXCL5,激活巨噬细胞CXCR2,诱导其向SPP1+免疫抑制表型极化,导致T细胞排斥。为克服YAP1抑制剂维替泊芬的脱靶毒性,研究开发了巨噬细胞膜仿生中空介孔二氧化硅纳米平台(M@O-VNPs),共载维替泊芬与奥沙利铂,实现肿瘤选择性YAP1抑制及免疫原性细胞死亡诱导,重塑免疫微环境,显著增强PD-1单抗疗效。该策略为克服胃癌免疫治疗抵抗提供了全新方案。
今天给大家解读一篇3月发表在《Cell Reports Medicine》上的题目为“Enhancing gastric cancer immunotherapy: Insights from multi-omics analysis and innovations in photodynamic-chemotherapy nanoplatforms.”的文章。本研究旨在克服胃癌免疫检查点阻断疗法的耐药性。首先,通过对接受新辅助ICB治疗的胃癌患者组织进行多组学分析,鉴定出肿瘤细胞固有转录因子YAP1是驱动免疫抑制微环境和治疗耐药的关键。进而,为了安全有效地靶向YAP1,研究构建了巨噬细胞膜伪装的中空介孔二氧化硅纳米颗粒M@O-VNPs,用于共递送YAP1抑制剂verteporfin和化疗药奥沙利铂。临床前研究表明,该纳米平台能够靶向肿瘤、抑制YAP1信号轴、诱导强大的免疫原性细胞死亡,从而将免疫抑制微环境逆转为免疫激活状态,显著增强抗PD-1疗法的疗效。研究为胃癌免疫治疗耐药提供了新的机制见解和一种有前景的组合治疗策略。(请持续关注我们,每天为您解读最新见刊的文献!)想薅生信资料羊毛?直接在对话框回复 “资料”,免费领取干货大礼包!包括数据集、绘图代码、图表复现、思路总结、参考文献……0代码!鼠标点点点即可轻松完成5-10分生信SCI全文复现!
不想做实验,没数据,还想要快速发表文章,没问题的!公共数据库就是我们的数据宝藏!没思路不用担心,作为专业的生信团队,我们很乐意为你们效劳,提供研究路线设计和数据挖掘分析,扫码联系我们吧!



团队成员合影(位于上海陆家嘴中心,可随时预约参观)
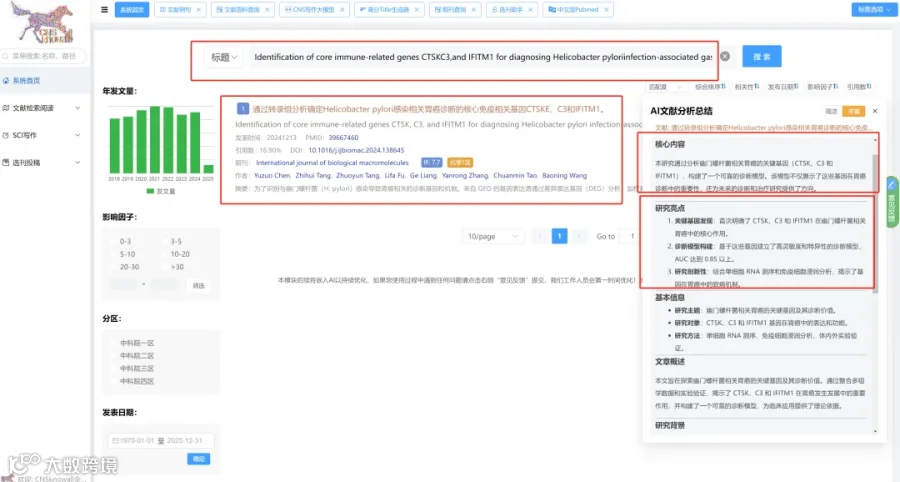
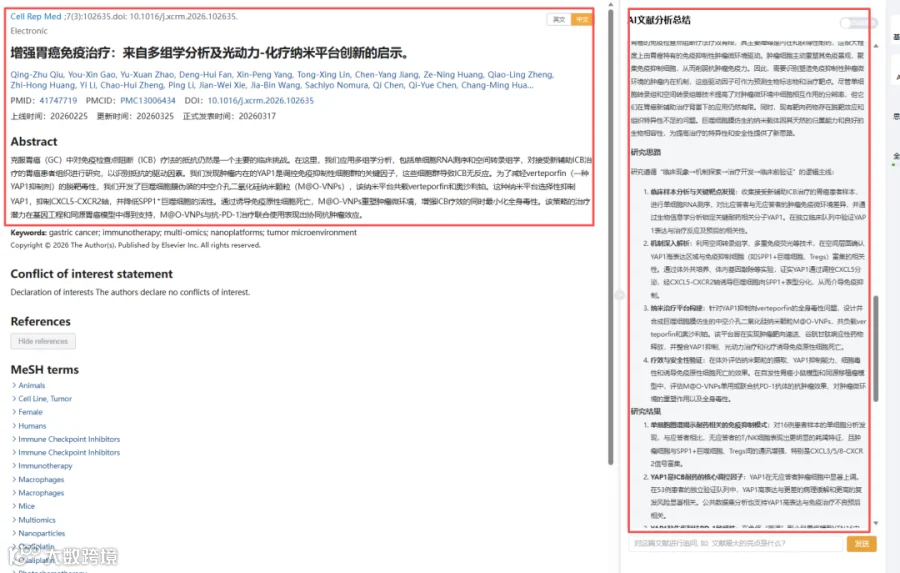
题目:《增强胃癌免疫治疗:来自多组学分析及光动力-化疗纳米平台创新的启示》Enhancing gastric cancer immunotherapy: Insights from multi-omics analysis and innovations in photodynamic-chemotherapy nanoplatforms
发表期刊:Cell Reports Medicine
影响因子:10.6
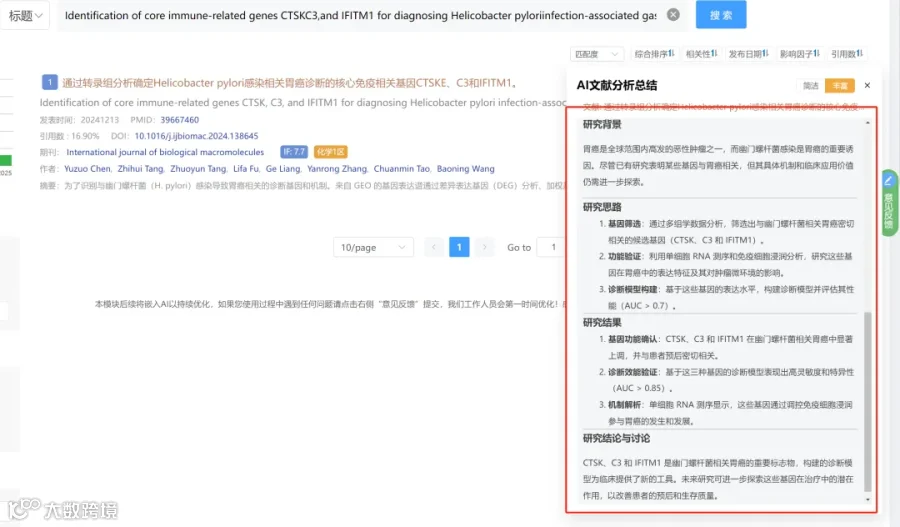
研究背景:
胃癌的免疫检查点阻断疗法疗效有限,其主要障碍是内在和获得性耐药,这很大程度上由胃癌特有的免疫抑制性肿瘤微环境驱动。肿瘤细胞主动重塑其免疫景观,聚集免疫抑制细胞,从而削弱抗肿瘤免疫力。因此,需要识别塑造免疫抑制性肿瘤微环境的肿瘤内在机制,这些驱动因子可作为预测生物标志物和治疗靶点。尽管单细胞转录组和空间转录组等技术提高了对肿瘤微环境中细胞相互作用的分辨率,但它们在胃癌新辅助治疗背景下的应用仍然有限。同时,现有靶向药物存在脱靶效应和组织特异性不足的问题。巨噬细胞膜仿生的纳米载体因其天然的归巢能力和良好的生物相容性,为提高治疗的特异性和安全性提供了新思路。
CNSknowall 平台 Pubmed+AI 快速提炼全文要点
研究思路:
研究遵循“临床现象→机制探索→治疗开发→临床前验证”的逻辑主线:
- 临床样本分析与关键靶点发现
收集接受新辅助ICB治疗的胃癌患者样本,进行单细胞RNA测序,对比应答者与无应答者的肿瘤免疫微环境差异,并通过生物信息学分析锁定关键耐药相关分子YAP1。在独立临床队列中验证YAP1表达与治疗反应及预后的相关性。
- 机制深入解析
利用空间转录组学、多重免疫荧光等技术,在空间层面确认YAP1高表达区域与免疫抑制细胞(如SPP1+巨噬细胞、Tregs)富集的相关性。通过体外共培养、体内基因敲除等实验,证实YAP1通过调控CXCL5分泌,经CXCL5-CXCR2轴诱导巨噬细胞向SPP1+表型分化,从而介导免疫抑制。
- 纳米治疗平台构建
针对YAP1抑制剂verteporfin的全身毒性问题,设计并合成巨噬细胞膜仿生的中空介孔二氧化硅纳米颗粒M@O-VNPs,共负载verteporfin和奥沙利铂。该平台旨在实现肿瘤靶向递送、谷胱甘肽响应性药物释放,并整合YAP1抑制、光动力治疗和化疗诱导免疫原性细胞死亡。
- 疗效与安全性验证
在体外评估纳米颗粒的摄取、YAP1抑制能力、细胞毒性和诱导免疫原性细胞死亡的效果。在自发性胃癌小鼠模型和同源移植瘤模型中,评估M@O-VNPs单用或联合抗PD-1抗体的抗肿瘤效果、对肿瘤微环境的重塑作用以及全身毒性。
研究亮点:
- 肿瘤固有的YAP1与胃癌新辅助ICB耐药相关
YAP1在无应答者的肿瘤细胞中显著上调,其高表达与较差的免疫治疗反应和生存预后相关。
- YAP1通过CXCL5-CXCR2-SPP1巨噬细胞轴促进免疫抑制
YAP1激活促进肿瘤细胞分泌CXCL5,通过作用于巨噬细胞上的CXCR2受体,诱导其分化为免疫抑制性的SPP1+表型。
- 巨噬细胞膜纳米平台实现肿瘤选择性YAP1抑制
构建的M@O-VNPs利用巨噬细胞膜的天然归巢能力增强肿瘤靶向性,并在肿瘤微环境的高谷胱甘肽条件下响应性释放药物。
- 联合靶向YAP1和PD-1阻断增强体内抗肿瘤免疫
在自发性及同源移植的胃癌模型中,光激活的M@O-VNPs与抗PD-1抗体联用,表现出协同抗肿瘤效应。
研究结果:
- 单细胞图谱揭示耐药相关的免疫抑制模式
对16例患者样本的单细胞分析发现,与应答者相比,无应答者的T/NK细胞表现出更明显的耗竭特征,且肿瘤细胞与SPP1+巨噬细胞、Tregs间的通讯增强,特别是CXCL3/5/8-CXCR2信号富集。
- YAP1是ICB耐药的核心调控因子
YAP1在无应答者肿瘤细胞中显著上调。在53例患者的独立验证队列中,YAP1高表达与更差的病理缓解和更高的复发风险显著相关。公共数据集分析也支持YAP1高表达与免疫治疗不良预后相关。
- YAP1缺失恢复抗PD-1敏感性
在免疫“荒漠”型小鼠胃癌模型YTN16中,敲除Yap1可显著抑制肿瘤生长,并使其对抗PD-1治疗变得敏感。
- 多组学揭示YAP1相关的免疫空间架构
空间转录组分析显示,YAP1高表达区域富集SPP1+巨噬细胞和Tregs,而YAP1低表达区域富集CD8+和CD4+ T细胞。大样本免疫组化证实了YAP1表达与免疫细胞浸润的负相关关系。
- 阐明YAP1-CXCL5-CXCR2-SPP1作用轴
肿瘤细胞YAP1表达与CXCL5分泌正相关。体外外实验证明,YAP1过表达通过CXCL5-CXCR2信号轴诱导巨噬细胞SPP1表达;阻断该轴可逆转此效应。
- 验证YAP1抑制剂verteporfin的疗效与毒性
在自发性胃癌小鼠模型中,verteporfin能抑制肿瘤生长并下调YAP1,但引起了肠道黏膜损伤和脾脏免疫细胞凋亡等全身毒性。
- M@O-VNPs纳米平台的构建与表征
成功合成并表征了M@O-VNPs,其具有约56 nm的粒径、巨噬细胞膜蛋白涂层、以及在模拟肿瘤微环境条件下谷胱甘肽响应性的药物释放特性。
- M@O-VNPs在体外实现靶向YAP1抑制与免疫原性细胞死亡诱导
纳米颗粒被胃癌细胞高效内吞,选择性抑制YAP1,并在光激活下产生ROS,与奥沙利铂协同诱导强烈的免疫原性细胞死亡,释放HMGB1、ATP,并促进钙网蛋白暴露。
- M@O-VNPs在体内重塑肿瘤免疫微环境
在荷瘤小鼠中,M@O-VNPs能靶向蓄积于肿瘤,光激活后显著抑制肿瘤内YAP1,诱导特异性强的免疫原性细胞死亡,增加效应T细胞和成熟树突状细胞浸润,减少Tregs和SPP1+巨噬细胞,并将巨噬细胞向M1表型极化。
- M@O-VNPs与抗PD-1的协同抗肿瘤效应
在自发性及移植瘤胃癌模型中,光激活的M@O-VNPs联合抗PD-1治疗,相比任一单药或游离药物组合,能更显著地抑制肿瘤生长、促进肿瘤细胞凋亡,且未引起明显的全身毒性或体重下降。
研究总结:
本研究确定YAP1是胃癌新辅助ICB治疗耐药的关键肿瘤内在驱动因子,其通过CXCL5-CXCR2轴促进SPP1+免疫抑制性巨噬细胞积累。为克服靶向YAP1的毒性挑战,研究开发了巨噬细胞膜仿生的M@O-VNPs纳米平台。该平台能够实现肿瘤选择性YAP1抑制,并协同诱导免疫原性细胞死亡,从而有效重塑肿瘤免疫微环境,将“冷”肿瘤转化为“热”肿瘤,最终显著增强抗PD-1疗法的疗效。
讨论指出,YAP1除本研究发现的功能外,可能还通过其他途径介导免疫逃逸。本研究开发的纳米平台为临床转化提供了一个概念框架,未来或可通过使用自体巨噬细胞膜或合成仿生膜进行优化。YAP1表达可通过免疫组化在临床样本中便捷检测,使其有望成为预测生物标志物,YAP1高表达患者或可从该联合策略中获益。研究的局限性包括样本来源于接受免疫化疗联合治疗的患者,以及需要在前瞻性临床研究中进一步验证该信号轴的预测价值。
结果译文:
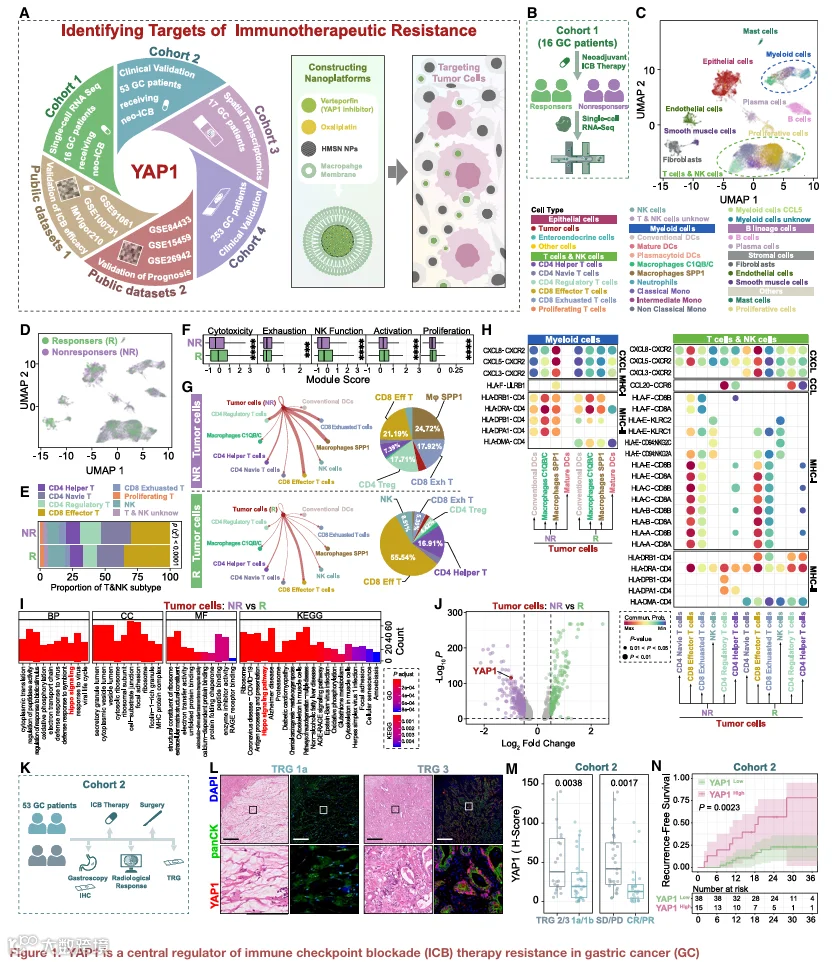
1.新辅助ICB应答者与非应答者的单细胞景观揭示免疫抑制性通讯模式
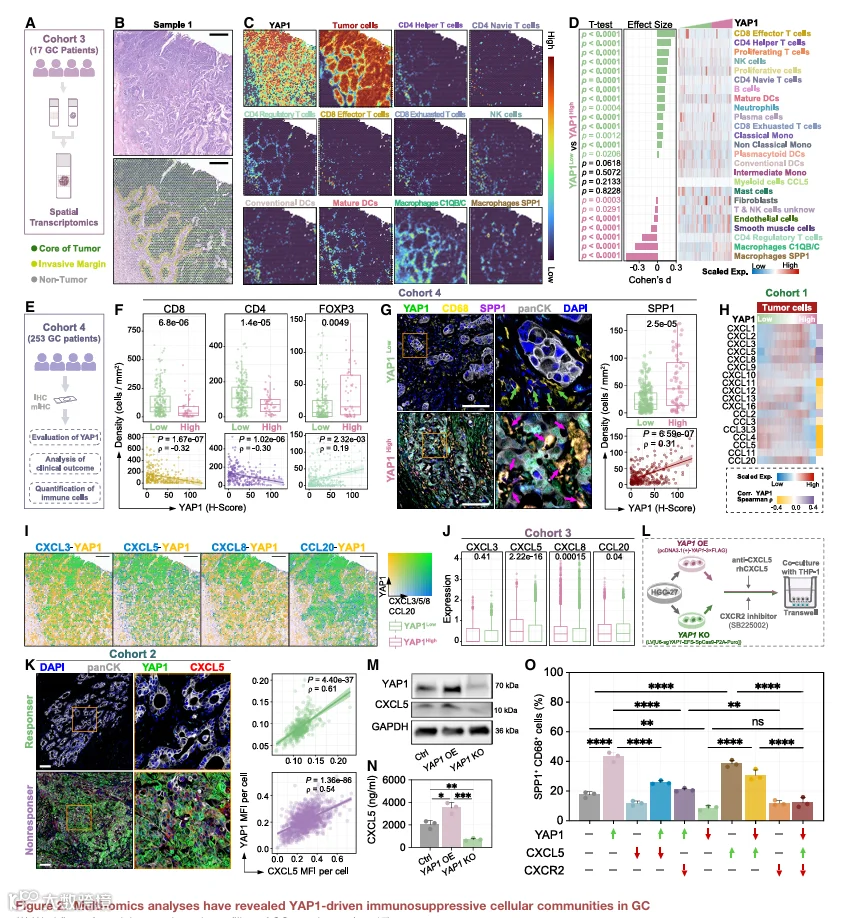
为探究新辅助免疫治疗抵抗的机制,我们收集了16例接受新辅助免疫检查点阻断治疗的胃癌新鲜肿瘤样本,用于scRNA-seq(图1B)。经质量控制和批次校正后,保留了45,929个细胞的转录组图谱,并将其分为29个转录上不同的簇(图1C和S1)。InferCNV分析进一步将上皮区室内恶性瘤细胞与非恶性上皮及肠内分泌细胞区分开来(图S2A-S2E)。
应答者与非应答者之间的比较揭示了细胞组成和免疫活性的显著差异(图1D-1F)。应答者的T细胞和自然杀伤(NK)细胞表现出增强的效应和增殖程序,而非应答者则表现出显性的耗竭特征。主要免疫亚群在两组间的分布差异显著(χ²检验,p<0.0001)。CellChat分析揭示了不同的肿瘤-免疫通讯景观(图1G)。在应答者中,肿瘤细胞主要与CD8+效应T细胞和CD4+辅助T细胞相互作用,与活跃的免疫参与一致。相反,非应答者显示向SPP1+巨噬细胞、调节性T细胞(Tregs)和耗竭CD8+ T细胞的信号增强,表明免疫抑制性通讯转变。配体-受体分析显示,应答者中肿瘤-T细胞相互作用主要由MHC I类和II类通路主导,而非应答者中肿瘤细胞与SPP1+巨噬细胞之间的CXCL3/5/8-CXCR2信号富集。此外,非应答者中肿瘤-Treg通讯显著涉及CCL20-CCR6轴,支持协同免疫抑制信号网络的出现。
2.YAP1相关转录和临床特征与免疫治疗无应答相关
为进一步探究非应答者免疫抑制表型的内在分子特征,我们比较了NR和R患者肿瘤细胞之间的差异表达基因(DEGs),并进行了GO和KEGG富集分析(图1l和1J)。非应答者中多条信号通路显著激活,包括Hippo信号级联。值得注意的是,其核心调控分子YAP1在NR肿瘤细胞中显著上调。鉴于YAP1在肿瘤进展、干性和治疗抵抗中的已知作用,但其在免疫调节方面的表征有限,我们接下来将分析重点放在YAP1上,作为与免疫治疗无应答相关的潜在介质。
为描绘YAP1表达异质性,根据YAP1阳性肿瘤细胞的比例将患者分为YAP1High和YAP1Low组(详见STAR Methods),并将这些标签映射回单细胞数据(图S2G和S2H)。YAP1High患者中约90%的肿瘤细胞来自非应答者(图S2I),表明YAP1激活定义了与免疫治疗抵抗相关的主导转录表型。
这些发现的临床相关性在一个独立的53例接受新辅助ICB治疗的GC患者队列(队列2;图1K和S3A)中进行了评估。基于肿瘤退缩分级和影像学反应,93%的YAP1High患者被归类为非应答者(图1L和S3B)。YAP1在非应答者中的表达显著高于应答者,跨多个临床分层均如此(TRG1a/1b vs. TRG2/3:2.55倍;CR/PR vs. SD/PD:2.77倍;图1M),单变量和多变量分析均证实其与ICB无应答相关(图S3C)。YAP1High患者还表现出更高的3年复发率和降低的总生存期(图1N和S3D)。在三个独立的免疫治疗队列(IMvigor210、GSE91061和GSE100797)中观察到一致的趋势,YAP1表达升高与不良反应和较短生存期相关(图S3D和S3E)。
综合这些发现,将YAP1定义为与免疫治疗无应答相关的肿瘤内在转录特征,并支持进一步研究其在免疫逃逸中的机制作用。
3.YAP1缺失在免疫荒漠型GC模型中恢复抗PD-1敏感性
为进一步在体内测试YAP1对抗PD-1治疗的功能影响,我们采用了小鼠GC细胞系YTN16,该细胞系呈现免疫荒漠表型和对PD-1阻断的内在抵抗。YTN16细胞被改造以建立稳定的Yap1敲除系,对照和Yap1-KO细胞均皮下植入C57BL/6小鼠(图S4A)。一旦肿瘤达到150-200mm³,小鼠接受14天的抗PD-1治疗。如预期,PD-1阻断在对照肿瘤中未产生显著肿瘤退缩(图S4B-S4D)。相比之下,Yap1缺失显著削弱了肿瘤生长,且重要的是,添加抗PD-1治疗使Yap1-KO肿瘤体积比抗PD-1处理的对照肿瘤减少3.4倍(p=0.0079)。一致地,肿瘤重量显著下降,Yap1-KO+抗PD-1组表现出最显著的组织学退缩和凋亡,如H&E和TUNEL染色所示(图S4E)。这些体内发现功能性地验证了YAP1是PD-1抵抗的肿瘤内在决定因素,支持其作为免疫治疗无应答机制性贡献者的作用。
为进一步阐明肿瘤-免疫相互作用的空间特征,我们对17例接受胃切除术患者的肿瘤标本进行了ST测序(队列3;图2A)。利用H&E染色结合基因表达数据,将肿瘤区域与邻近非肿瘤区域区分开来(图2B)。然后根据YAP1表达中位数将肿瘤斑点分为YAP1High和YAP1Low组。应用稳健细胞类型分解(RCTD)算法整合scRNA-seq和ST数据,推断每个空间斑点内的细胞类型组成(图2C和S5)。识别出不同的免疫结构,CD8+效应T细胞和CD4+辅助T细胞富集于YAP1Low区域,而SPP1+巨噬细胞、C1QB+/C+巨噬细胞和FOXP3+ Tregs在YAP1High区域占优势(图2D)。多重免疫荧光染色证实了这些空间模式,显示YAP1Low肿瘤巢内有密集的CD8+GZMB+效应T细胞和非Treg CD4+ T细胞浸润,而FOXP3+ Tregs则优先定位于YAP1High肿瘤细胞附近(图S6)。在一个更大的包含253例GC样本的免疫组织化学队列(队列4)中,免疫浸润定量显示YAP1Low肿瘤中CD8+和CD4+ T细胞密度分别增加2.14倍和1.6倍,同时YAP1High病例中SPP1+巨噬细胞升高2.31倍(图2E和2G)。
这些空间分析将肿瘤YAP1表达与以效应T细胞浸润减少和SPP1表达巨噬细胞积累为特征的免疫抑制微环境联系起来。
5.肿瘤YAP1与巨噬细胞SPP1通过CXCL5-CXCR2轴的旁分泌串扰
鉴于SPP1+巨噬细胞在治疗抵抗中的免疫抑制作用,我们检测了肿瘤源性YAP1与巨噬细胞之间的潜在串扰。配体-受体分析表明,非应答者中肿瘤-巨噬细胞通讯富集了CXCL3/5/8-CXCR2信号(图1H)。在肿瘤细胞中,YAP1表达与CXCL5(p=0.3508,p<0.0001)和CXCL8(p=0.3045,p<0.0001)的相关性最强(图2H)。空间转录组学显示YAP1与CXCL3、CXCL5、CXCL8和CCL20的定位一致(图2I),其中CXCL5表达在YAP1High区域比YAP1Low区域高1.42倍(p<0.0001,图2J)。在蛋白水平上,ICB治疗肿瘤样本的多重免疫荧光显示非应答者中YAP1和CXCL5高表达且空间共定位强,而应答者中两种标志物均以显著较低水平表达,同时保持相似的空间模式(图2K)。相比之下,CXCL3和CXCL8未表现出与YAP1相当的空间关联(图S7A),支持CXCL5是GC中主要的YAP1相关趋化因子。
在巨噬细胞方面,CXCR2几乎由所有SPP1+巨噬细胞表达,尽管并非所有CXCR2+细胞都表达SPP1(图S7B)。为功能性测试这一轴,我们构建了YAP1过表达和YAP1敲除的HGC27细胞(图2L)。CXCL5蛋白水平与YAP1表达相关,经western blotting和ELISA证实(图2M和2N)。在共培养实验中,肿瘤YAP1过表达使巨噬细胞SPP1表达增加2.45倍(p<0.0001),而YAP1缺失则产生相反效应(图2O和S7C)。这种诱导作用被CXCL5中和抗体或CXCR2抑制所消除,重组CXCL5在YAP1敲除条件下恢复了SPP1表达,该作用再次被CXCR2抑制所阻断。
与这些体外发现一致,使用YTN16对照或Yap1敲除肿瘤(给予重组CXCL5、CXCL5中和抗体或CXCR2抑制剂SB225002)的体内分析显示,YAP1缺失显著降低了SPP1+巨噬细胞比例。这一降低被重组CXCL5逆转,并被CXCR2抑制消除(图S8A-S8D)。综合这些数据表明,肿瘤内在YAP1通过CXCL5-CXCR2依赖机制促进GC中巨噬细胞SPP1的诱导。
上述发现提示YAP1驱动的肿瘤-巨噬细胞信号传导有助于免疫抑制性TME的形成,促使我们研究其在GC中的临床相关性。鉴于高度微卫星不稳定性(MSI-H)和Epstein-Barr病毒(EBV)阳性状态是公认的免疫治疗反应预测因子,进一步评估YAP1在这些亚组中的表达是必要的。然而,由于MSI-H和EBV阳性的低发生率,需要更大的队列。因此,我们分析了队列4,一个具有长期随访数据的良好注释GC队列(n=253)。YAP1表达在MSI-H和EBV阳性肿瘤中显著低于MSI-L/MSS和EBV阴性病例(图S9A),支持其与免疫治疗反应性的潜在预测相关性。生存分析显示,YAP1高表达患者的5年总生存率为34%,而YAP1低表达患者为58%(图S9B)。多变量分析进一步确定YAP1为GC的独立负面预后因素(图S9C)。这一趋势在三个公共数据集(GSE26942、GSE15459和GSE84433)中得到验证,其中YAP1低表达患者始终表现出更优的5年生存率(图S9D)。
总之,这些数据将YAP1上调与不良临床结局联系起来,支持其作为GC免疫治疗抵抗和预后的生物标志物潜力。
7.经典YAP1抑制剂维替泊芬阻断肿瘤生长但伴有显著副作用
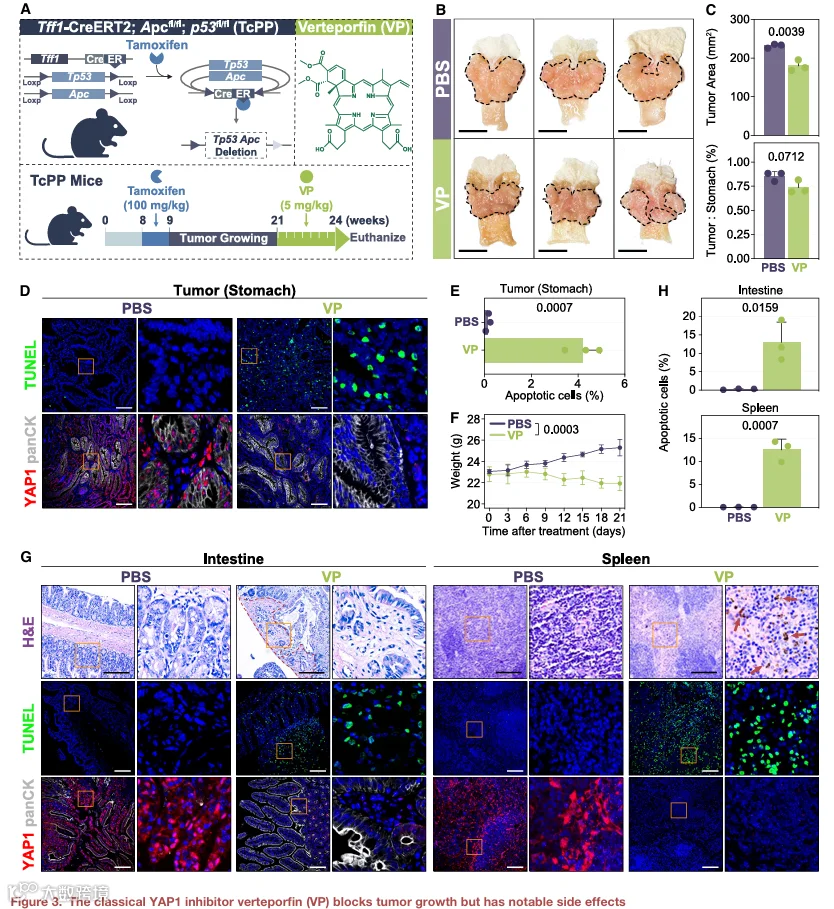
鉴于靶向YAP1在免疫治疗抵抗背景下的潜在治疗相关性,并考虑到体内基因操作的现实局限性,我们接下来评估了药理学YAP1抑制的抗肿瘤效应。VP,一种经典的YAP1抑制剂,可破坏YAP1-TEAD结合,被用作药理学工具评估YAP1抑制的体内后果。为此,我们构建了Tff1-CreERT2-Apcf/f,p53f/f(TcPP)小鼠模型,这是一种条件性敲除模型,通过靶向胃上皮中的Tff1、Apc和Tp53诱导自发性GC(图3A)。这种自发性肿瘤模型提供了比皮下肿瘤模型更符合生理的TME和更强大的药物评估平台。
在12周的tamoxifen给药后,对TcPP小鼠进行药物干预,VP通过尾静脉注射,剂量为1mg/kg,每2天一次,持续3周。PBS注射对照组用于比较(图3A)。VP治疗导致肿瘤缩小(p=0.0039;图3B和3C),并伴随胃肿瘤组织中YAP1抑制和部分肿瘤细胞凋亡(p=0.0007;图3D和3E)。然而,在治疗期间,小鼠体重适度减轻(p=0.0003;图3F),提示潜在不良反应。值得注意的是,肠黏膜损伤伴随YAP1抑制(肠:p=0.0159;脾:p=0.0007;图3G和3H)。YAP1对维持肠黏膜稳态和再生至关重要,其缺乏可能损害受损肠组织的修复。此外,在VP处理小鼠的脾脏中检测到铁蛋白沉积和免疫细胞显著凋亡,表明可能存在脾脏毒性(图3G和3H)。
总之,尽管VP有效抑制了肿瘤生长,但其副作用相当大。除了观察到的肠道和脾脏毒性外,YAP1抑制还可能导致肝毒性、肺纤维化和肾功能损害,因其在器官稳态中的关键作用。此外,VP的内在光敏性构成另一个治疗挑战——其光触发活性可诱导全身光毒性。然而,若利用得当,这一特性也提供了治疗机会,通过活性氧(ROS)生成实现光动力肿瘤消融。这种独特的光动力能力使VP区别于其他YAP1抑制剂,既是风险也是机制优势。因此,提高VP的选择性和可控激活对于将其双重YAP1抑制和光动力潜力转化为安全有效的治疗至关重要。
8.特异性靶向肿瘤内在YAP1的光动力-化疗纳米平台的开发与表征
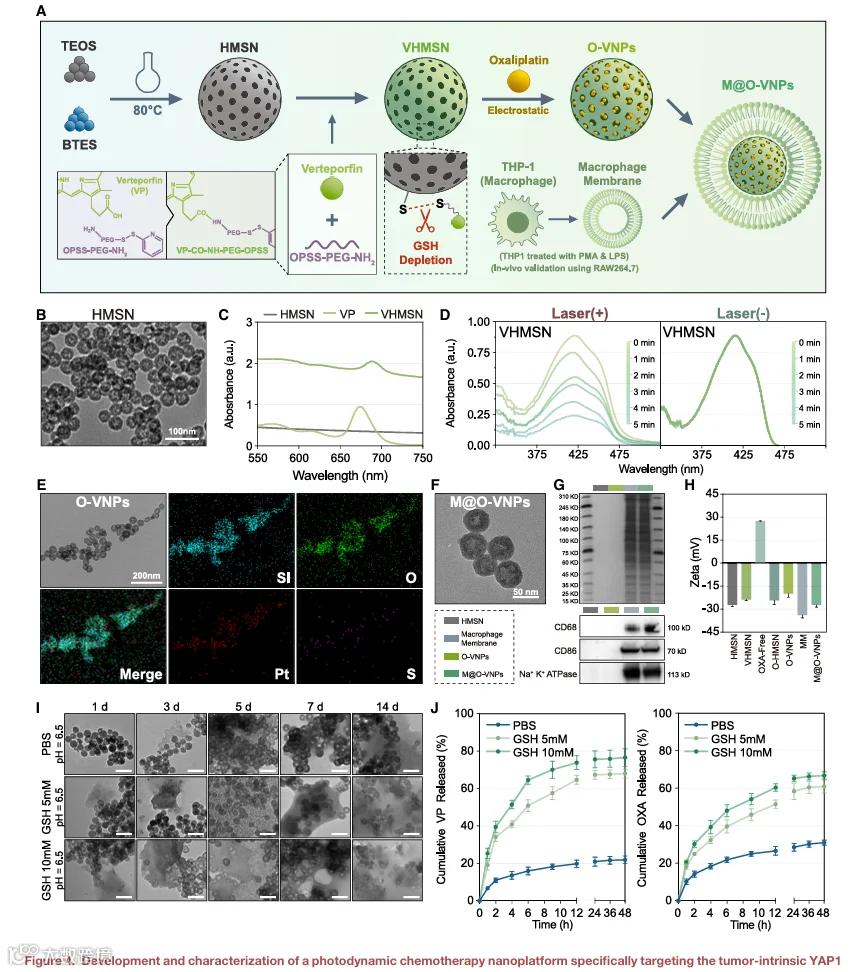
靶向递送系统对于降低VP的全身毒性和提高肿瘤选择性至关重要。巨噬细胞膜伪装的HMSNs提供了一种利用巨噬细胞固有肿瘤归巢特性的递送策略。这些纳米颗粒促进VP在肿瘤内优先积累,从而限制正常组织暴露。除了增强瘤内VP递送外,该平台还使VP能够双重用于YAP1抑制和光激活诱导免疫原性细胞死亡。
VP单独作为根除肿瘤的独立疗法是不充分的,尽管其潜力巨大,仍需联合治疗。奥沙利铂(OXA),一种铂类化疗药物,与VP表现出协同配对效应。OXA诱导免疫原性细胞死亡(ICD),并与VP产生的ROS协同作用以增强抗肿瘤功效。此外,YAP1抑制可增强铂类药物(如OXA)的有效性,使得该组合在克服化疗耐药方面尤为有力。
基于这些考虑,合成了共载VP和OXA的巨噬细胞膜伪装介孔二氧化硅纳米平台(M@O-VNPs)(图4A)。HMSNs采用BTES和TEOS生成的核壳结构制备,经氨蚀刻后得到平均直径约40nm的球形纳米颗粒。所得HMSNs具有高比表面积(364.89m²/g)和约3.78nm的介孔,能够实现高效药物装载(图S10A和S10B)。X射线衍射分析表明无定形二氧化硅结构(图S10C)。VP通过CDI化学与OPSS-PEG-NH₂偶联,经¹H NMR光谱确认(图S10D),随后连接至巯基化HMSNs形成具有谷胱甘肽响应释放特性的VP负载纳米颗粒(VHMSNs)。VHMSNs显示VP装载效率约9.37%,并保留了游离VP的特征吸收曲线(图4C)。在激光照射(690nm,100mW/cm²)下,VHMSNs在体外产生单线态氧,如DPBF降解所示,而无照射时未观察到变化(图4D)。OXA随后通过静电吸附封装于介孔结构内,得到O-VNPs,其装载量约16.73%,经能量色散谱确认(图4E)。然后将巨噬细胞膜包覆于O-VNPs上生成M@O-VNPs,最终直径约56nm(图4F)。SDS-PAGE和western blot分析确认关键巨噬细胞膜蛋白(包括CD68和CD86)得以保留(图4G),zeta电位测量进一步验证了膜的成功包覆(图4H)。
所得纳米平台表现出谷胱甘肽响应性降解,能够在还原性TME内实现可控药物释放(图4I)。升高的细胞内GSH水平可裂解连接VP与载体的二硫键,从而促进VP的快速和选择性释放,同时促进OXA从介孔基质中释放(图4J)。一致地,药物释放实验显示在生理条件下渗漏极少,而在富含GSH的环境中释放显著加速(图S10E)。鉴于肿瘤组织的高还原潜力,该设计支持VP和OXA的优先瘤内递送,实现有效的YAP1抑制和光激活诱导免疫原性细胞死亡。
9.光激活M@O-VNPs实现肿瘤选择性YAP1抑制和ICD诱导
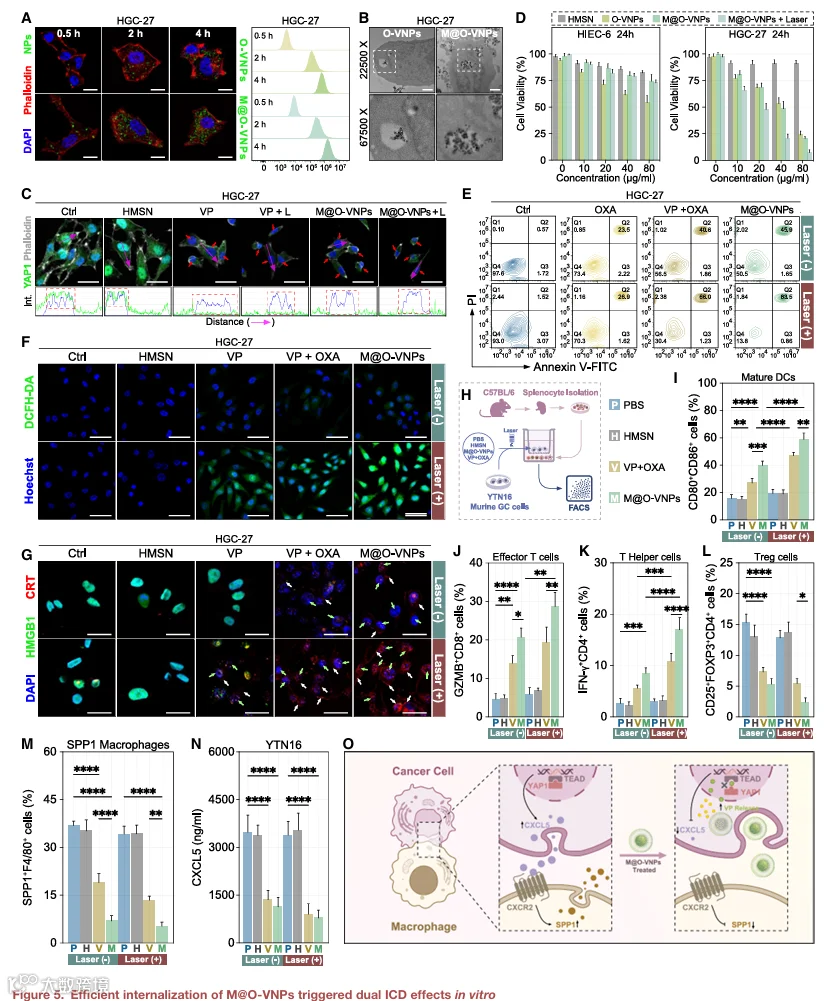
为评估细胞摄取,O-VNPs和M@O-VNPs用异硫氰酸荧光素(FITC)标记,并通过共聚焦激光扫描显微镜(CLSM)、流式细胞术(FACS)和透射电子显微镜(TEM)进行分析。与未包覆的O-VNPs相比,M@O-VNPs在HGC-27 GC细胞中显示显著增强的内化(图5A和5B)。相比之下,巨噬细胞膜包覆后小鼠脾细胞的摄取显著减少(图S11A),表明有效的免疫逃避和网状内皮系统吞噬清除减少。这种仿生表面修饰有望延长循环时间并促进肿瘤优先积累。
内化后,细胞内谷胱甘肽裂解M@O-VNPs内的二硫键,导致VP释放并随后抑制YAP1-TEAD结合(图5C和S11B),该效应独立于激光照射。采用CCK-8实验评估细胞毒性和生物安全性。M@O-VNPs对人正常肠上皮细胞(HIEC-6)影响极小,即使在激光照射后仍保持超过70%的细胞活力(图5D)。相比之下,光激活M@O-VNPs将HGC-27细胞活力降至10%以下。Annexin V-FITC/PI染色进一步显示M@O-VNPs诱导的凋亡率高于游离VP-OXA,且激光照射后进一步增强(图5E)。一致地,DCFH-DA实验证实光激活后产生强劲ROS(图5F和S11C)。
光激活VP通过ROS生成诱导ICD,而OXA作为一种免疫原性药物也可引发ICD。我们假设VP和OXA的组合将增强ICD反应。我们评估了经典ICD标志物,包括HMGBl释放、钙网蛋白(CRT)转位和ATP分泌。如预期,共处理VP和OXA后观察到HMGBl和ATP释放水平升高,且光激活后效应更显著(图5G、S11D和S11E)。同样,游离VP-OXA和M@O-VNP处理细胞在激光照射后均检测到细胞表面CRT暴露(图5G和S11D),作为抗原呈递细胞(APCs)的“吃我”信号。
这些结果证实光激活M@O-VNPs实现选择性肿瘤细胞摄取,并通过YAP1抑制和ROS介导的细胞毒性有效诱导免疫原性细胞死亡。
10.光激活M@O-VNPs重编程YAP1-CXCL5-SPP1轴并重塑肿瘤免疫微环境
为更好地模拟复杂免疫微环境,将小鼠脾细胞随后与经相同处理条件的YTN16 GC细胞共培养(图5H)。FACS分析显示,光激活M@O-VNPs引发最强劲的免疫激活,特征为成熟树突状细胞(mDCs)、CD8+效应T细胞和IFN-γ+CD4+ T辅助细胞频率升高,同时Treg细胞和SPP1+巨噬细胞显著减少(图5I-M和S12)。值得注意的是,两种基于VP的治疗,尤其是M@O-VNPs,显著抑制了CXCL5分泌并破坏了巨噬细胞中的CXCL5-CXCR2信号,从而减弱SPP1表达(图5N)。综合这些发现表明,光激活M@O-VNPs不仅增强ICD并提升抗原呈递,还通过破坏YAP1-CXCL5-SPP1轴重编程肿瘤-免疫界面(图5O),最终培育免疫许可微环境。
鉴于SPP1+肿瘤相关巨噬细胞(TAMs)与常规M2样巨噬细胞共享若干转录特征和功能标志物,我们接下来检测了M@O-VNPs是否影响巨噬细胞极化。对与处理肿瘤细胞共培养的THP-1源性巨噬细胞进行FACS分析显示,其表型向M1样亚型转变,同时M2相关标志物减少(图S13A-S13D)。细胞因子分析进一步证实了这一趋势(图S13E和S13F)。该效应可能由YAP1抑制结合双重ICD激活诱导的损伤相关分子模式(DAMPs)释放所驱动。
尽管M@O-VNPs在体外表现出强大的靶向和抗肿瘤活性,但转化应用前需进行全面的体内安全性评估。因此,我们在野生型C57BL/6小鼠中进行了毒理学评估。溶血实验显示该纳米平台具有良好的血液相容性(图S14A)。小鼠静脉注射M@O-VNPs,剂量为5、10、20或40mg/kg,每2天一次,持续28天(图S14B)。接受5或10mg/kg的动物维持稳定体重,与PBS对照相当,而20和40mg/kg组观察到显著体重减轻(图S14C)。血清生化显示高剂量组从第4天起肝损伤标志物(ALB、ALT和AST)升高,H&E染色证实组织病理学肝损伤(图S14D和S14E)。在20和40mg/kg组还观察到额外的肠道和脾脏损伤,以及剂量依赖性死亡率。相比之下,10mg/kg或更低剂量处理的小鼠显示正常体重、保留的肝肾功能,且无明显组织学异常。因此,选择10mg/kg作为后续体内研究的安全剂量。
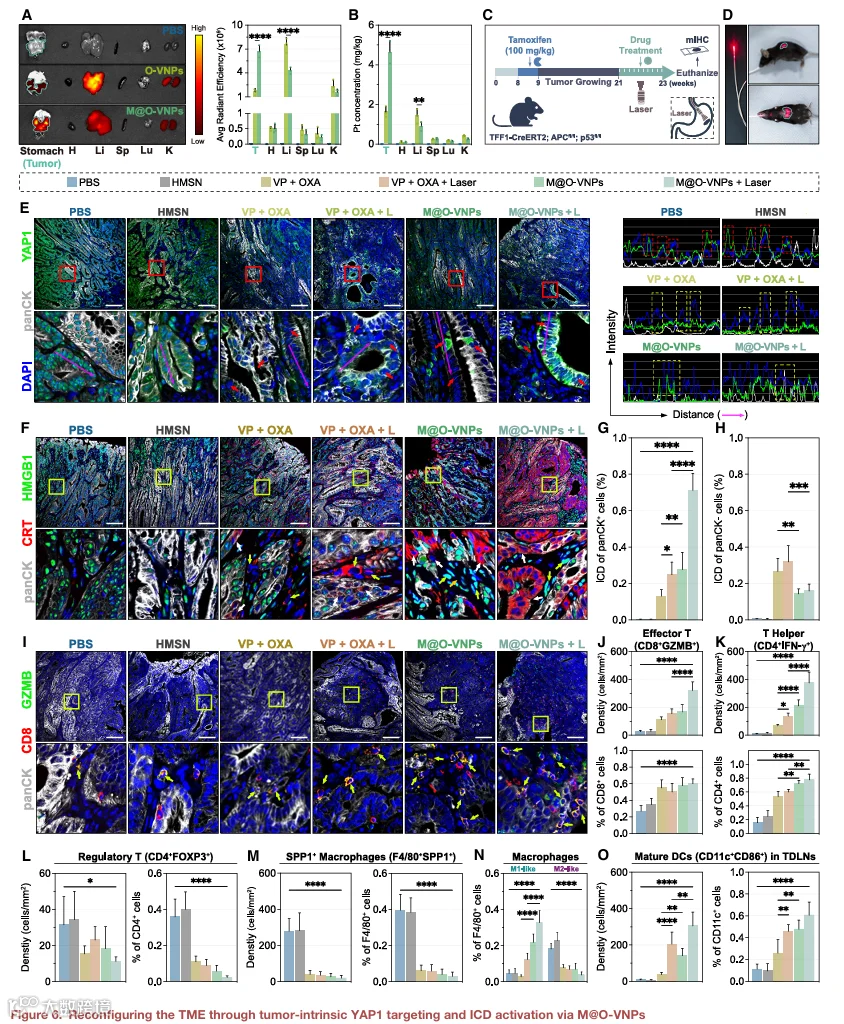
为评估体内肿瘤靶向效率,将ICG标记的纳米颗粒静脉注射至携带自发性胃肿瘤的TcPP小鼠中。注射后24小时,收集主要器官进行离体IVIS成像,并通过ICP-MS定量铂(Pt)生物分布。如图6A和6B所示,M@O-VNPs显示在胃肿瘤内优先积累,与未包覆的O-VNPs相比肝脏信号显著降低。定量分析显示肿瘤荧光增加4.1倍,肝脏信号降低43%。一致地,ICP-MS分析证实瘤内Pt水平增加2.8倍,而肝脏Pt滞留降至O-VNP组的63%。这些结果证明了体内巨噬细胞膜包覆赋予的增强肿瘤选择性和减少脱靶积累。
12.M@O-VNPs通过靶向肿瘤源性YAP1重塑TME
鉴于M@O-VNPs的精准靶向能力,我们预期治疗后TME将发生显著调节。如图6C所示,纳米平台给药后24小时施加激光照射。为有效照射胃部,将定制设计的扩散发射器插入食管(图6D)。治疗2周后,处死小鼠,将肿瘤连续切片以分析TME内的变化。
肿瘤切片的多色荧光成像显示,游离VP-OXA和M@O-VNPs均有效抑制了YAP1的核转位。值得注意的是,M@O-VNPs对非肿瘤性YAP1表达的影响减弱(图6E)。与游离VP形成鲜明对比,M@O-VNPs对非靶组织(如肠道和脾脏)中YAP1表达的影响可忽略不计(图S15A和S15B)。
对ICD标志物的进一步分析证实,光激活M@O-VNP处理的肿瘤表现出最强劲的ICD反应,与体外观察一致(图6F)。值得注意的是,M@O-VNPs诱导的ICD反应高度选择性,主要靶向肿瘤细胞。相比之下,游离VP-OXA引发了更弥漫、非特异性的ICD效应(图6G和6H)。这些结果突显了光激活M@O-VNPs触发的ICD反应的卓越效力和特异性。
为进一步探究M@O-VNPs对TME的体内影响,我们进行了免疫细胞分析,重点研究先前显示与YAP1活性强相关的细胞群体及ICD相关免疫相互作用中关键涉及的细胞群体。光激活M@O-VNP处理后,观察到肿瘤内效应T细胞(CD8+GZMB+)浸润显著增加,与PBS组相比增加12.02倍,与“VP+OXA+L+”组相比增加2.10倍。效应T细胞占总CD8+ T细胞的比例也分别增加2.23倍和1.2倍。这种免疫激活模式在T辅助细胞(CD4+IFN-γ+)中得到体现(图6I-6K和S16A),表明强劲的免疫刺激效应。相反,免疫抑制性调节性T细胞(CD4+FOXP3+)和SPP1+巨噬细胞(F4/80+SPP1+)群体在M@O-VNP处理后分别显著下降至PBS组的35.78%和7.20%(图6L、6M、S16B和S16C)。这一转变与共培养结果一致,观察到肿瘤组织中M2样巨噬细胞显著减少,伴随M1样巨噬细胞增加(图6N和S16D)。此外,在胃周肿瘤引流淋巴结(TDLNs)中,光激活M@O-VNP处理后检测到成熟和迁移DC(CD11c+CD86+)显著增加,与PBS组相比增加39倍,与“VP+OXA+L+”组相比增加1.50倍。这种增强的DC活性导致细胞毒性T细胞的强劲激活,促进有效肿瘤细胞杀伤(图6O和S16E)。
总之,M@O-VNPs成功靶向胃肿瘤细胞,导致YAP1抑制和ICD诱导。YAP1阻断和ICD诱导的联合效应在光激活M@O-VNP处理后使TME转变为免疫激活状态。
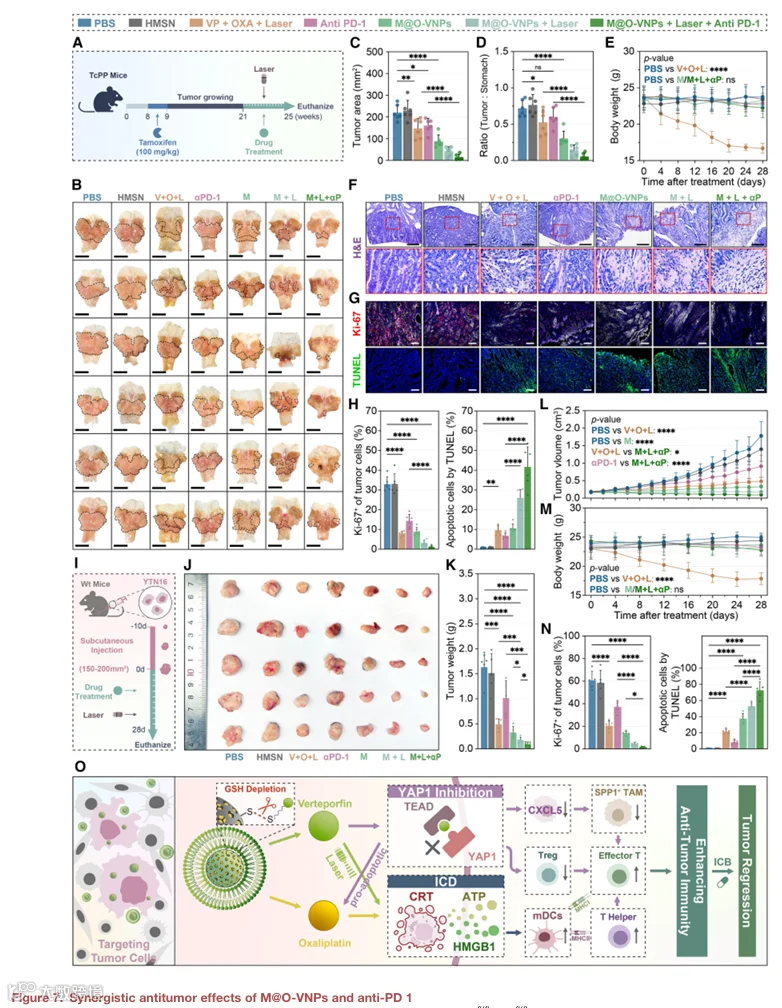
13.M@O-VNPs与抗PD-1的协同抗肿瘤效应
对于成功的免疫治疗,免疫激活的TME是先决条件,而M@O-VNPs可有效重塑这种环境。基于这一原理,我们接下来评估了M@O-VNPs联合PD-1阻断的抗肿瘤疗效(图7A)。治疗4周后,处死TcPP小鼠进行肿瘤和器官分析。胃肿瘤面积相对于总胃大小的定量显示,光激活M@O-VNP处理后肿瘤显著退缩,与PBS对照相比减少5.27倍,与“VP+OXA+L+”组相比减少3.53倍(图7B-7D)。重要的是,将光激活M@O-VNPs与PD-1抑制联合进一步增强了肿瘤退缩,与PD-1单药治疗相比实现10.66倍的减少。体重监测表明M@O-VNP处理的小鼠波动极小,而接受光激活游离VP加OXA的小鼠则表现出显著体重减轻,与全身毒性一致(图7E)。组织病理学分析证实“M@O-VNP+laser+”组肿瘤退缩显著,这在“M@O-VNP+laser++anti-PD-1”组中最为明显(图7F)。一致地,Ki-67染色显示增殖指数在M@O-VNP+laser和联合组中分别降至抗PD-1组的22.71%和7.23%,而TUNEL实验显示凋亡相应增加(图7G和7H)。值得注意的是,在M@O-VNP处理的小鼠中未观察到主要器官(包括肠道和脾脏)的明显组织学损伤,与游离VP+OXA组观察到的器官毒性形成对比(图S17)。
为进一步验证治疗稳健性,我们将这些研究扩展至YTN16同系模型(图7I)。治疗4周后,光激活M@O-VNPs联合PD-1阻断使肿瘤体积与PD-1单药治疗相比减少9.61倍,与“VP+OXA+laser”组相比减少10.79倍(图7J-7L),且未影响体重(图7M)。组织学评估证实肿瘤显著退缩(图S18A)。免疫荧光分析进一步显示,与两个对照治疗组相比,Ki67阳性细胞显著减少,凋亡肿瘤细胞相应增加(图7N、S18B和S18C)。
总之,这些结果表明巨噬细胞膜伪装增强了M@O-VNPs的肿瘤选择性和治疗效果,以最小全身毒性实现有效肿瘤控制。其与PD-1阻断的联合在互补体内模型中进一步放大抗肿瘤反应,支持克服GC免疫治疗抵抗的联合策略。
更多结果和补充图表:doi: 10.1016/j.xcrm.2026.102635
长按二维码关注我们,用最短的时间和最高的效率学习更多数据分析方法!
扫描上方二维码或登录平台官网后添加CNSknowall客服微信咨询!官网地址:
https://cnsknowall.com
CNSknowall:24年最新问世的遥遥领先的科研数据(0代码生信+统计学)分析平台,同时含有机制图模块+汉化版Pubmed融合Deepseek高效筛选目标文献+SCI文献例句/语料检索模块+OPenAI官方GPT接口,>500款CNS级别图表皆可一秒内一键出图,登录即秒变数据分析大神,体验前所未有的便捷数据分析之旅,开启科研天骄之路!
可向下滑动批阅!