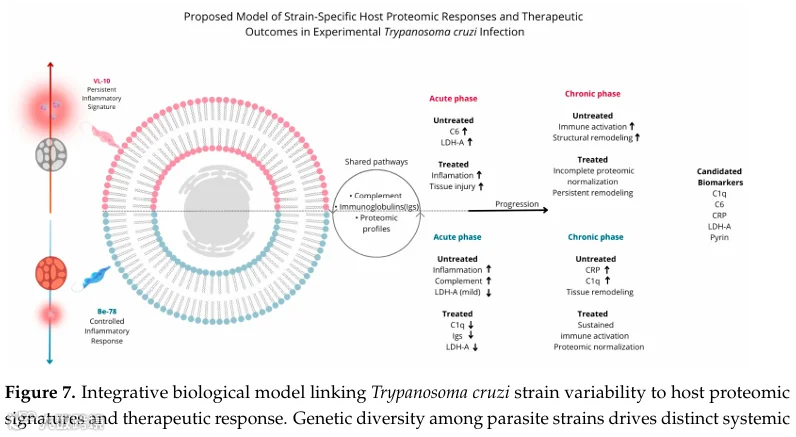
同一种药,为何有的恰加斯病患者有效,有的却无效?答案藏在感染虫株的遗传密码里。这项研究利用非标记定量血清蛋白质组学,对比了苯并咪唑敏感株Be-78和耐药株VL-10感染小鼠在急慢性期的血清蛋白谱。结果惊人:敏感株感染后,补体C1q、免疫球蛋白、LDH-A等炎症和损伤标志物在治疗后几乎完全恢复正常;而耐药株感染后,C6、Pyrin炎性小体、肌营养不良蛋白、伴肌动蛋白等持续飙升,即使治疗也难以逆转。这项研究首次从血清蛋白质组层面证实,虫株遗传异质性是决定疗效的核心因素,并鉴定出一套可区分治疗反应的血清标志物组合。
今天给大家解读一篇3月发表在《Microorganisms》上的题目为“Serum Proteomics Provides Novel Biomarkers of Inflammation, Tissue Injury, and Therapeutic Response in Experimental Chagas Disease.”的文章。本研究利用定量无标记UHPLC-MS/MS蛋白质组学技术,分析了感染BNZ敏感(Be-78)和耐药(VL-10)克氏锥虫株的小鼠在急性和慢性阶段的血清蛋白质组。结果发现,两种菌株诱导了截然不同的蛋白质组学特征。敏感株感染后,BNZ治疗能使大多数失调蛋白(如补体、免疫球蛋白、炎症介质)恢复正常,显示出高效的治疗响应。相反,耐药株感染后,即使经过治疗,补体(特别是C6)、细胞损伤标志物(LDH-A)和结构蛋白(肌养蛋白、伴肌动蛋白)仍持续高水平,提示更严重的组织损伤和持续的炎症状态。研究最终鉴定出一组与菌株毒力、炎症负担和药物疗效相关的关键蛋白,为恰加斯病的生物标志物开发和治疗监测提供了新见解。(请持续关注我们,每天为您解读最新见刊的文献!)想薅生信资料羊毛?直接在对话框回复 “资料”,免费领取干货大礼包!包括数据集、绘图代码、图表复现、思路总结、参考文献……0代码!鼠标点点点即可轻松完成5-10分生信SCI全文复现!
不想做实验,没数据,还想要快速发表文章,没问题的!公共数据库就是我们的数据宝藏!没思路不用担心,作为专业的生信团队,我们很乐意为你们效劳,提供研究路线设计和数据挖掘分析,扫码联系我们吧!



团队成员合影(位于上海陆家嘴中心,可随时预约参观)
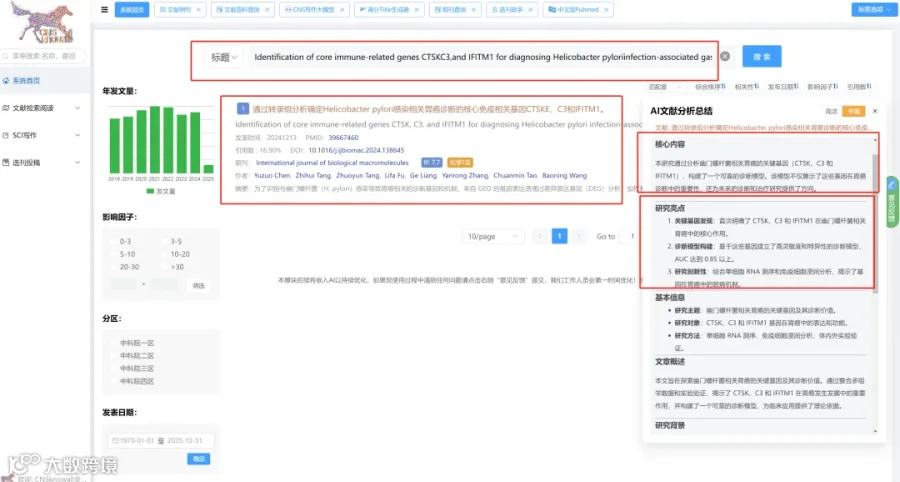
题目:《血清蛋白质组学为实验性恰加斯病的炎症、组织损伤和治疗反应提供了新的生物标志物》Serum Proteomics Provides Novel Biomarkers of Inflammation, Tissue Injury, and Therapeutic Response in Experimental Chagas Disease
发表期刊:Microorganisms
影响因子:4.2
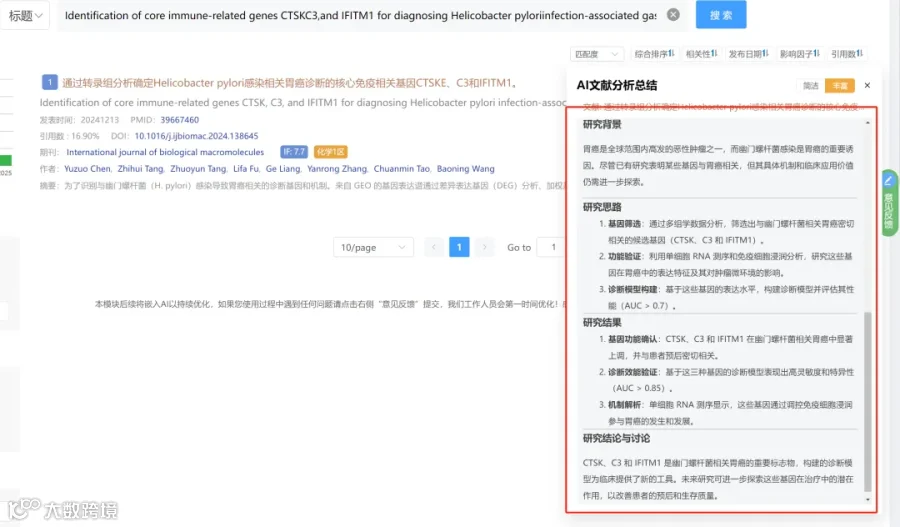
研究背景:
恰加斯病仍是重要的公共卫生挑战,由原生动物克氏锥虫引起,全球约600-700万人感染,其中30-40%会发展为慢性心脏或消化系统疾病。目前缺乏能够准确预测临床进展或监测治疗反应(特别是在慢性期)的可靠血清生物标志物。克氏锥虫的遗传变异性(不同离散分型单元DTU)是影响临床异质性、毒力、组织趋向性、炎症反应和药物疗效(如BNZ)的主要因素之一。虽然急性期治疗有效,但慢性期疗效有限且缺乏可靠评估标志物。已有研究表明,宿主-寄生虫相互作用深刻调节补体、免疫球蛋白、氧化应激、能量代谢和结构重塑等通路。蛋白质组学分析是揭示疾病严重程度、炎症强度和治疗结果分子通路的有力工具,但大多数研究缺乏同时评估不同菌株、不同感染阶段和药物直接效应的整合方法。因此,存在一个关键空白:缺乏能够表征克氏锥虫遗传变异性和药物治疗如何在整个感染过程中重塑血清蛋白质组的研究。本研究正是为填补此空白而设计。
CNSknowall 平台 Pubmed+AI 快速提炼全文要点
研究思路:
- 动物模型与实验设计
使用雌性瑞士小鼠,分别感染BNZ敏感株(Be-78)和BNZ耐药株(VL-10)。每个菌株的感染又分为急性期和慢性期两个阶段,每个阶段设置三个实验组:未感染对照组(CN)、感染未治疗组(NT20)、感染后BNZ治疗20天组(T20)。
- 样本采集与处理
在实验结束时采集血清,使用BCA法进行蛋白定量。对每个实验组随机选取3份血清样本进行蛋白质组学分析,样本不混合。
- 蛋白质组学分析
采用定量无标记技术。流程包括:血清蛋白经还原、烷基化、胰蛋白酶酶解后,通过UHPLC-MS/MS(高分辨质谱)进行数据依赖性采集。原始数据用PEAKS Studio软件进行蛋白鉴定和定量,以UniProt小鼠数据库为参考,FDR≤1%。
- 生物信息学分析
在PEAKS中进行归一化和差异蛋白筛选。通路分析使用PANTHER v18.0软件。通过比较不同组别(CN vs. NT20 vs. T20)及不同菌株间的差异表达蛋白,分析菌株特性、感染阶段和治疗对血清蛋白质组的影响。
研究亮点:
- 发现新型生物标志物
筛选出C1q、C6、C反应蛋白、LDH-A、Pyrin、肌养蛋白、伴肌动蛋白、内收蛋白和肌球蛋白等作为组织损伤、持续炎症和治疗反应的潜在血清生物标志物。
- 揭示菌株依赖性差异
明确显示克氏锥虫的遗传变异性是决定系统性炎症、组织损伤和对BNZ治疗响应性的主要因素。敏感株(Be-78)炎症可控且治疗后恢复,耐药株(VL-10)则表现为持续炎症和有限治疗反应。
- 构建整合模型
提出了一个整合生物学模型,将寄生虫菌株变异性与宿主蛋白质组特征及治疗反应联系起来,突出了寄生虫遗传多样性在疾病监测和改进诊断治疗策略中的重要性。
研究结果:
- 全局血清蛋白质组特征
鉴定到数百种蛋白,检测动态范围约五个数量级。感染后,血清白蛋白比例下降,炎症相关蛋白(如补体C3、转铁蛋白、血红素结合蛋白、妊娠区带蛋白PZP)升高,载脂蛋白A-I(ApoA1)降低。治疗(特别是对Be-78株)可部分或完全逆转这些变化。
- 菌株特异性蛋白组学特征
- Be-78敏感株
急性期NT20组有105个独有蛋白,主要为免疫球蛋白和经典补体成分,治疗后仅有7个独有蛋白,表明炎症大幅缓解。慢性期NT20组仍有32个独有蛋白(包括肌肉再生相关蛋白Tetra-nectin),治疗后降至5个,显示治疗有效但慢性期恢复不彻底。
- VL-10耐药株
急性期NT20组有53个独有蛋白,包括补体C6、细胞连接蛋白(Ezrin、Junction plakoglobin),治疗后仍有4个独有蛋白。慢性期NT20组有25个独有蛋白(包括结构蛋白Nebulin)。更关键的是,慢性期治疗组仍有30个独有蛋白(超过Be-78),提示治疗后血清蛋白质组仍高度失调,存在代偿或适应机制。
- 差异表达蛋白分析
- Be-78株
急性期NT20组中C1qA/B、多种免疫球蛋白、VCAM-1、组织蛋白酶S等显著上调,但乳酸脱氢酶A(LDH-A)和凝血因子XIII A链显著下调。治疗后(T20组),上述蛋白基本恢复正常。
- VL-10株
急性期NT20组中C6上调超过100倍,C1qA/B、组织蛋白酶S也显著升高,LDH-A显著升高(与Be-78相反)。治疗(T20组)仅能部分降低这些蛋白水平,远未恢复正常。慢性期NT20和T20组中Nebulin、肌养蛋白、肌球蛋白XVIIIb、α-内收蛋白等结构蛋白持续显著变化,Pyrin(炎症小体标志物)显著升高。
- 菌株间比较
两株共有的差异蛋白(如C1q、C6、免疫球蛋白)在VL-10株中的上调幅度远大于Be-78株。LDH-A在VL-10株中持续高表达,而在Be-78株中降低。BNZ治疗后,Be-78株的共有蛋白显著恢复,而VL-10株则保持高水平,直接反映了治疗耐药性。
研究总结:
- 主要结论
克氏锥虫的遗传变异性是决定系统性炎症、组织损伤和对BNZ疗法响应性的主要因素。BNZ敏感株感染后,治疗可带来广泛的蛋白质组恢复正常;而BNZ耐药株感染后,无论是在急性还是慢性期,均表现为炎症介质、补体、免疫球蛋白和结构蛋白的持续升高,与持续炎症和较低的治疗有效性一致。
- 生物标志物候选
C1q亚基、C6、C反应蛋白、LDH-A、Pyrin以及肌养蛋白、伴肌动蛋白、内收蛋白和肌球蛋白等结构蛋白,是组织损伤、持续炎症和治疗反应的有前景的血清生物标志物。特别是VL-10感染小鼠中这些标志物的不完全正常化,提示存在与治疗耐药性相关的独特、可测量的分子特征。
- 临床及研究意义
研究结果支持开发用于监测恰加斯病进展和治疗结果的多标志物血清面板。同时强调了将克氏锥虫遗传多样性纳入诊断工具和治疗策略设计中的重要性。该研究通过血清蛋白质组学揭示了与寄生虫毒力、炎症负担和药物疗效相关的生物学特征,为未来精准医疗提供了基础。
- 讨论与局限性
作者指出,使用非去白蛋白的血清虽然能提供更广泛的蛋白质组图谱,但可能限制对细胞因子或低分子量蛋白的检测。此外,研究采用的是小鼠模型,其结果向人类患者的转化需要进一步验证。尽管如此,该研究为恰加斯病的生物标志物发现和治疗监测提供了重要的、系统性的新见解。
结果译文:
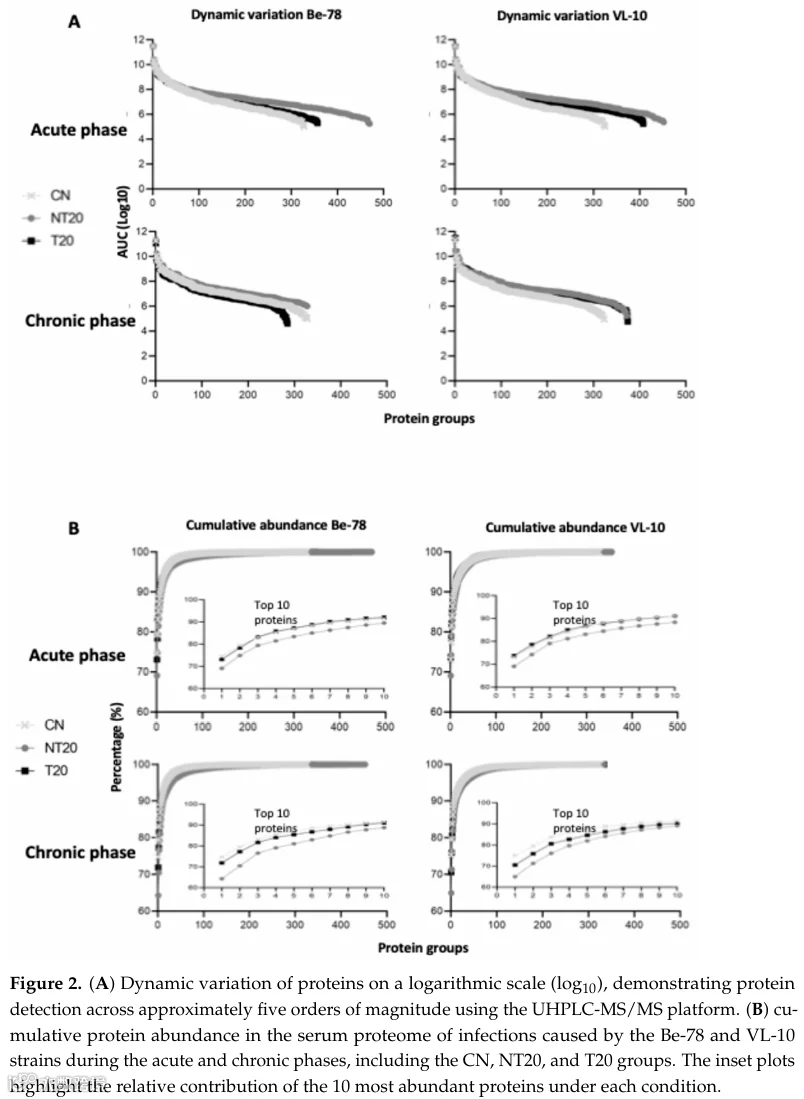
UHPLC-MS/MS方法在所有实验条件下均鉴定出一组稳健且可重复的血清蛋白。在急性期,Be-78虫株鉴定出517种蛋白,VL-10虫株鉴定出506种蛋白(表S1);在慢性期,分别鉴定出380和422种蛋白(表S2)。这些数量反映了良好的分析深度、适当的血清蛋白质组覆盖率以及样本和实验组间的技术一致性。完整的光谱采集指标——包括MS/MS扫描数、PSM、鉴定肽段数及每个蛋白的独特肽段分布——详见表1和表2。
全局丰度分布揭示了约五个数量级的动态范围(图2),这是非去除高丰度蛋白血清中鸟枪法分析的典型特征。样本中丰度最高的前10种蛋白被列出(表S3),其中白蛋白在所有条件下占总丰度的65-75%——这一模式在类似研究中被广泛记载。尽管白蛋白占主导,该方法仍能检测到大量生物学相关蛋白,突显了分析平台的强大性能。
在感染未处理组(NT20)中,观察到白蛋白比例降低,同时伴随炎症反应、铁/血红素代谢、补体系统和免疫球蛋白相关蛋白的显著增加——这一模式与克氏锥虫感染的病理生理学一致。其中,血清转铁蛋白因其在限制病原体铁利用中的重要作用而突出,血红素结合蛋白作为清除游离血红素和减轻氧化损伤的关键组分亦显著升高。这些蛋白的升高强化了感染期间铁/血红素代谢的深刻紊乱。
妊娠区带蛋白(PZP)也呈现显著增加,尤其在VL-10感染中。先前研究报道了其在实验性恰加斯病急性期的升高及其参与全身炎症过程,支持其作为潜在炎症标志物的相关性。
另一个突出组分是补体C3,在感染组中被鉴定为最丰富的蛋白之一。除在先天免疫中发挥核心作用外,C3还在慢性感染期间影响T细胞调节,突显其参与此处观察到的全身性反应。C3与多种免疫球蛋白的同步升高与克氏锥虫感染特征性的持续性免疫激活一致。
载脂蛋白A-I(ApoA1)在NT20组中显著降低,经BNZ治疗后部分或完全恢复,尤其在Be-78虫株中。该行为与ApoA1在脂质代谢和先天免疫中的作用一致。研究表明,克氏锥虫主要蛋白酶cruzipain可切割ApoA1,导致其在感染期间减少。此外,有研究将ApoA1确定为治疗反应的候选生物标志物,强化了本研究中观察到的模式一致性。
在慢性期,观察到α-1B-糖蛋白(A1BG)增加。尽管其功能尚未完全阐明,但已与通过与CRISP3相互作用调节炎症反应相关联。其升高可能反映了持续性炎症背景下晚期免疫适应机制。
综上,这些发现表明,尽管白蛋白是血清蛋白质组中的主导蛋白,但感染驱动了血清蛋白的深刻重塑,尤其是在NT20组中。与先天免疫、氧化应激、铁/血红素代谢、全身炎症和补体激活相关的通路被广泛调节。BNZ治疗逆转了部分改变,在Be-78虫株中更为显著,而VL-10表现出更轻微的调节,与其较低的药物敏感性一致。这些结果强化了克氏锥虫虫株内在差异和寄生虫控制程度直接影响血清蛋白质组的观点。
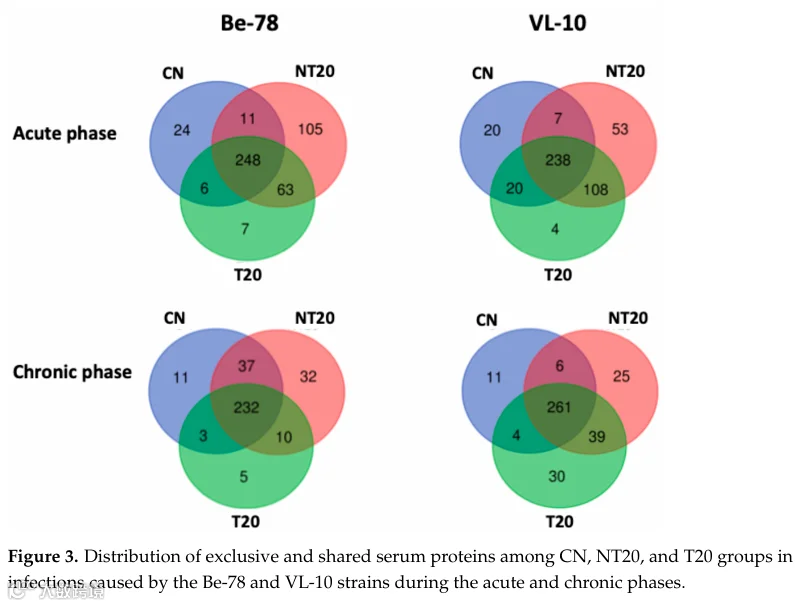
分析每种实验条件下独特检出的蛋白集合,揭示了急性期与慢性期之间以及Be-78与VL-10虫株之间的不同模式(图3;表S4)。这些谱图反映了炎症激活程度、免疫调节和组织重塑的差异——这些现象与感染的发病机制广泛相关。
在Be-78急性期,NT20组显示出最多的独特蛋白(105种),主要由免疫球蛋白和经典补体通路组分组成,与急性期特征性的体液免疫应答强烈激活一致。其中功能相关性最强的独特蛋白包括:糜蛋白酶样弹性蛋白酶2α(参与代谢机制和炎症反应);补体C2(经典补体通路的核心组分);以及簇集蛋白(片段),与炎症、凋亡和组织重塑过程相关。
感染Be-78虫株的T20组仅呈现7种独特蛋白,表明治疗后炎症活性降低和血清蛋白质组部分正常化,符合药物敏感虫株的预期行为。
在慢性期(Be-78感染),NT20组鉴定出32种独特蛋白,包括多种免疫球蛋白和补体调节因子,表明感染后很长时间仍存在持续性免疫激活——这是慢性恰加斯病中公认的现象。值得注意的蛋白包括:四连接素(与肌肉再生和组织重塑相关);补体因子H相关蛋白4(克氏锥虫常利用其进行免疫逃逸的替代补体通路调节因子)。另一方面,T20组仅呈现5种独特蛋白,强化了治疗后蛋白质组稳定的观点。
在VL-10虫株急性期,NT20组显示出53种独特蛋白,包括若干IgG链;补体C6(末端补体激活的经典标志物);以及埃兹蛋白、连接斑珠蛋白和MACF1等结构和细胞连接蛋白。该集合表明:先天和适应性免疫的强烈激活;以及对心脏完整性至关重要的蛋白质改变,与恰加斯心肌病中描述的结构性病变一致。
感染VL-10虫株的T20组仅呈现4种独特蛋白,表明在该耐药虫株中治疗对血清蛋白质组调节的影响有限。
在VL-10虫株感染慢性期,NT20组显示出25种独特蛋白,包括免疫球蛋白、补体调节因子和结构蛋白。值得注意的是,与肌原纤维组织和肌肉收缩性相关的伴肌动蛋白被鉴定出——常与慢性心脏病中的肌肉损伤相关联。有趣的是,T20组呈现出30种独特蛋白,数量高于Be-78观察到的数量,包括与肌肉收缩性、能量代谢和组织重塑相关的蛋白。该模式表明,即使在治疗后,VL-10虫株仍维持高度调节的血清蛋白质组,可能反映了代偿过程和感染后适应机制,与治疗反应性低的慢性模型观察结果一致。
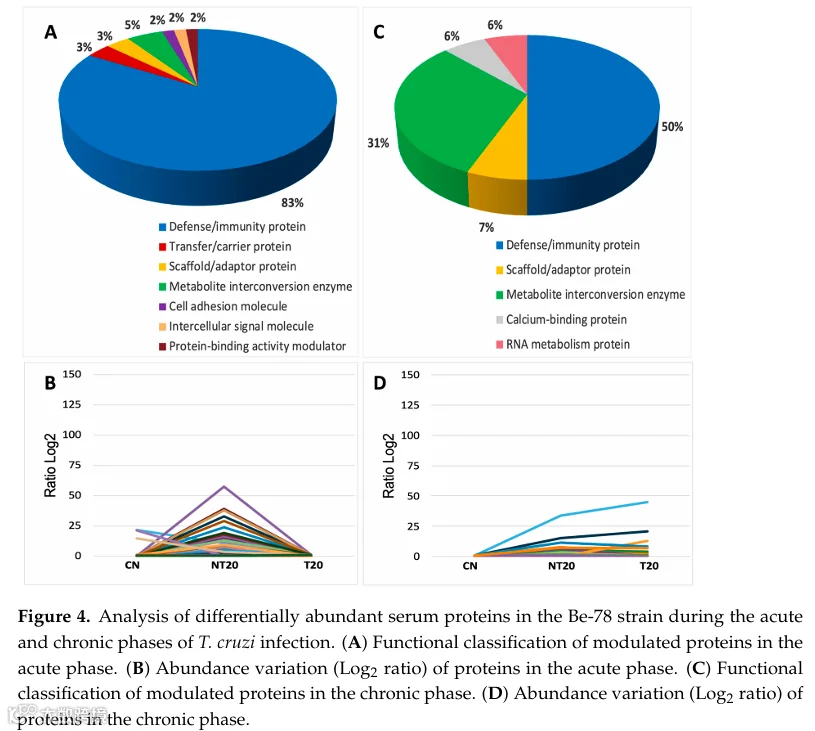
定量分析在Be-78虫株急性感染期鉴定出67种差异丰度蛋白(图S1和图4;表S5)。观察到的谱图反映了一种强烈且高度调节的全身炎症反应,以补体激活、免疫球蛋白和免疫激活相关介质为主导。
NT20组显示多种先天和适应性免疫组分的显著增加。在最受调节的蛋白中,C1qA和C1qB尤为突出(较CN高3-4倍),证实了经典补体通路的强烈激活。这一趋势与免疫球蛋白的大量扩增并行——超过50种重链、κ链和λ链免疫球蛋白呈现显著升高,其中许多的log2倍数变化超过15-30。该模式表明急性期高峰典型的旺盛体液反应,符合药物敏感模型。
参与免疫识别和调节的蛋白,如BPI折叠含家族A2、CD5抗原样蛋白和组织蛋白酶S,也显示出显著增加,反映炎症反应放大和抗原加工活性增强。此外,与黏附和细胞募集相关的蛋白如VCAM-1在NT20组中强烈升高,表明内皮激活,与恰加斯病急性期描述的病理生理学一致。
有趣的是,凝血相关蛋白表现出与强烈炎症状态下典型预期相反的行为。凝血因子XIII A链在NT20组中被完全抑制,提示炎症期间血凝块稳定性受损或过度消耗。同样,代谢标志物L-乳酸脱氢酶A链(LDH-A)在NT20中显著降低,在T20中完全消失。该结果可能反映负代谢调节、全身消耗增加或组织释放至血清减少,与经典的细胞损伤模式相反。
T20产生了血清蛋白质组的强力正常化。NT20中升高的大多数蛋白恢复至接近CN水平,包括免疫球蛋白、C1q和炎症介质。VCAM-1和α-烯醇酶等蛋白降至对照水平以下,表明治疗诱导的强效抗炎作用和代谢再平衡。这组发现清晰地证明了Be-78虫株的高治疗反应性,与其BNZ敏感表型一致。
总之,Be-78感染的急性期以补体和免疫球蛋白通路的强烈激活为特征,伴随炎症介质、抗原加工和免疫调节的强烈调节。此外,若干补体组分——包括C1qA和C1qB——显著升高,强化了Be-78虫株急性期特征的免疫高反应性。
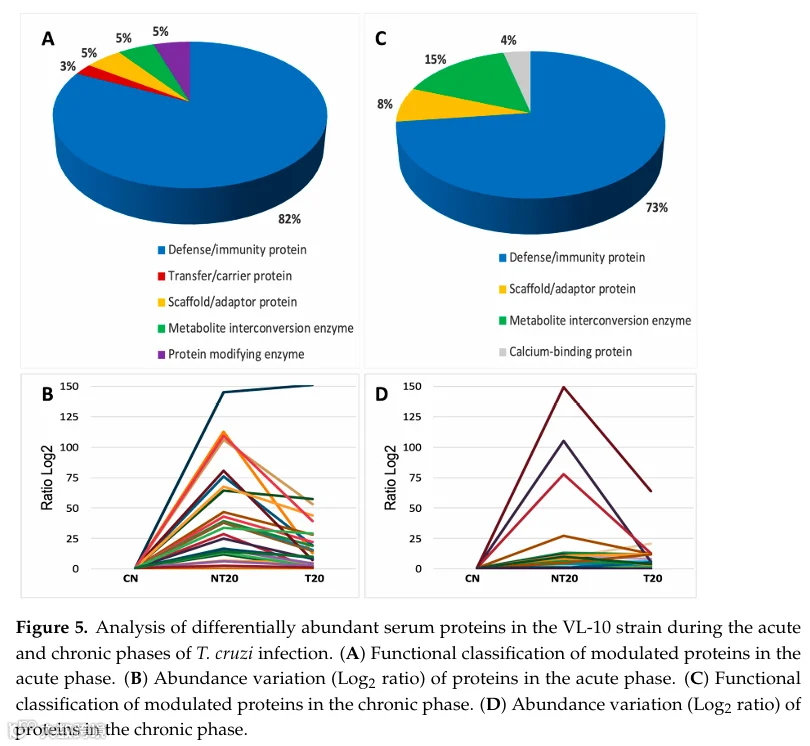
在VL-10虫株急性感染期,鉴定出53种差异丰度蛋白(图S2和图5;表S7)。与Be-78相比,VL-10的谱图揭示了一个更加复杂的炎症环境,表现为补体激活持续增强、组织损伤标志物显著以及BNZ治疗后仅部分消退。
在NT20组中,若干先天免疫组分显著升高。其中,末端补体通路组分C6的增加极为突出,达到极高值,表明强烈的膜攻击复合物形成和细胞溶解潜能。经典通路组分C1qA和C1qB也显著上调,尽管幅度略低于Be-78。这种补体特征表明VL-10虫株激发了一种剧烈且可能破坏性更强的体液免疫反应。
免疫球蛋白也呈现显著调节,重链、κ链和λ链水平升高,证实了B细胞应答的参与。然而,与Be-78相比,VL-10中差异丰度免疫球蛋白的总数较少,表明尽管存在强烈的体液激活,但克隆扩增模式不同或受到更严格的调控。其他炎症介质如BPI折叠含蛋白和组织蛋白酶S也显著增加,反映了VL-10虫株急性期特征的抗原加工机制、免疫识别和炎症信号放大。
组织损伤模式在该虫株中尤为明显。结构蛋白和与细胞完整性相关的蛋白被调节,包括细胞骨架蛋白片段和细胞动力学调节因子。与敏感虫株相比,该模式表明更严重的结构损害。白蛋白耗竭和低分子量炎症蛋白水平升高也加剧了这种更高致病侵袭性的情景。
T20仅诱导大多数受调节蛋白的部分降低。尽管C6、C1qA/B和组织蛋白酶S观察到持续下降,但其水平仍远高于CN,表明治疗无法完全逆转炎症状态。该行为与VL-10虫株的天然BNZ耐药表型一致。
差异丰度免疫球蛋白集合——尽管数量上少于Be-78——在NT20中显示中度升高,强化了体液应答的参与,尽管模式不那么旺盛。这可能反映了两虫株间炎症动态和抗原血症谱的内在差异。
总之,VL-10感染的急性期以补体过度激活、组织损伤标志物显著、炎症反应加剧和治疗调节有限为特征,形成与高毒力和低BNZ敏感性一致的蛋白质组学谱。与Be-78的对比突显了寄生虫遗传变异在塑造血清蛋白质组和决定治疗效力中的重要性。
在VL-10感染的慢性期,差异丰度蛋白揭示了持续性免疫激活、组织重塑和结构改变的持续情景,与该虫株的耐药表型一致(表S8)。观察到的模式表明,即使在长期感染后,动物仍维持高度调节的血清蛋白质组,提示残留炎症、持续心脏重塑和寄生虫控制不完全。
在NT20组中,观察到若干重链、κ链和λ链免疫球蛋白的持续增加,反映了耐药克氏锥虫模型慢性特征性的体液应答维持。此外,补体系统组分的调节蛋白,包括C1qA和替代通路蛋白片段,保持升高,表明持续的免疫激活。
VL-10感染慢性期的一个引人注目的方面是结构和肌原纤维蛋白的调节。其中,伴肌动蛋白作为肌动蛋白细丝组织和肌原纤维完整性所必需的蛋白脱颖而出。其在NT20组中的增加提示持续性肌肉损伤或晚期再生机制的激活,这些现象已在慢性心肌病背景下被描述。参与肌肉结构、能量代谢和收缩性的其他蛋白也被调节,强化了即使在感染晚期仍存在持续组织重塑的假设。
在治疗组(T20)中,观察到若干炎症和结构蛋白的部分降低;然而,差异丰度蛋白的总数及其变异幅度与Be-78虫株相比仍然很高。值得注意的是,VL-10 T20组展示的独特蛋白数量比慢性Be-78本身更多,表明即使在BNZ治疗后,这些动物的血清蛋白质组仍然广泛改变。该行为表明治疗不足以恢复蛋白质组稳态,使与炎症、免疫应答和结构重组相关的通路保持激活。
其他相关蛋白包括参与能量代谢的分子(如肌酸激酶亚型)、细胞外基质蛋白、细胞应激调节因子和钙结合蛋白,它们共同反映了与耐药虫株相关的慢性心肌病典型的病理生理适应。
总之,VL-10感染的慢性期以持续性免疫激活、持续的结构和代谢损伤、连续的组织重塑和有限的治疗调节为特征。这些发现强化了BNZ耐药性和寄生虫遗传变异对疾病进展和全身蛋白质组谱的影响。
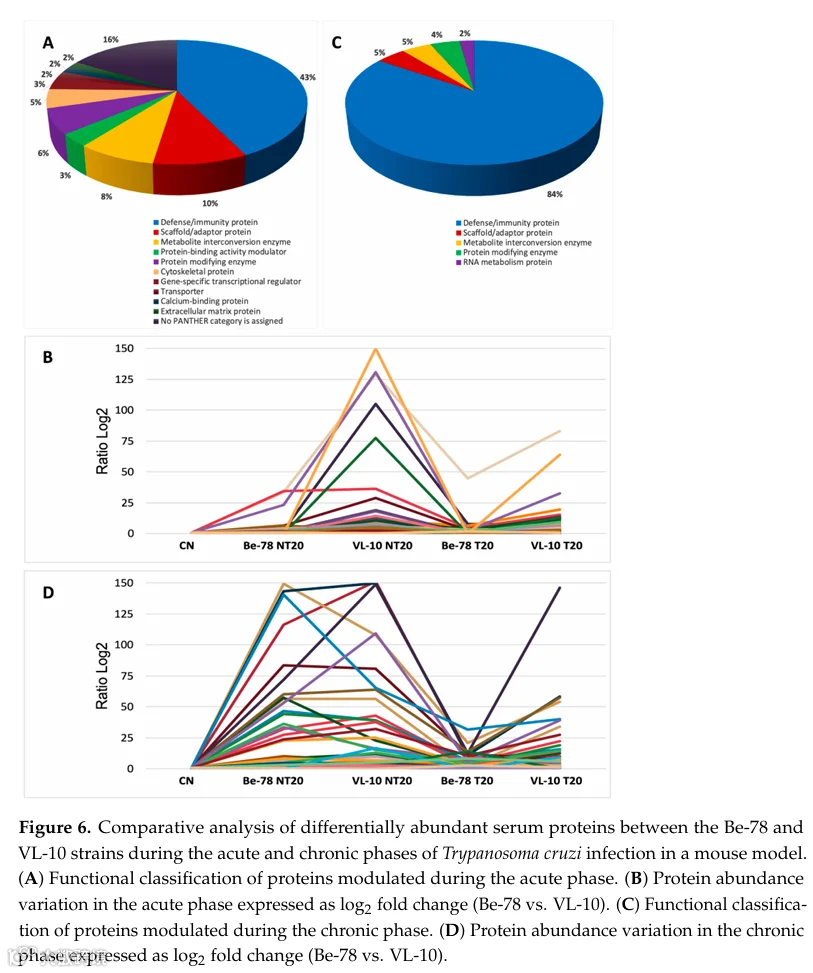
在急性感染期,鉴定出51种蛋白为Be-78和VL-10虫株所共有(图S3和图6;表S9),约占差异丰度蛋白总数的10%。该集合主要由补体系统组分、重链、κ链和λ链免疫球蛋白以及炎症蛋白组成,形成两种感染共同的全身炎症核心。
在共有蛋白中,C1qA、C1qB和C6尤为突出,表明两种虫株中经典和末端补体通路的强烈激活,尽管在VL-10中幅度更大,如该虫株中C6水平显著更高所示。共有的炎症和免疫调节蛋白——包括BPI折叠含蛋白、组织蛋白酶S、CD5抗原样蛋白和组织相容性2 Q区基因座4——提示与识别、加工和抗原呈递相关的通路同步激活。
代谢和全身反应标志物如血红素结合蛋白、磷脂转移蛋白和三甲基鸟苷合酶也在两种虫株中持续检测到,表明脂质代谢和血红素/铁转运的调整构成共有炎症轴的一部分。然而,并非所有共有蛋白表现出相同方向或强度的调节:例如,载脂蛋白C-III在若干组中显示轻微降低而非增加,而凝血因子XIII A链和纤连蛋白等蛋白在虫株和条件间表现出不同行为。
细胞损伤标志物L-乳酸脱氢酶A链在虫株间呈现不对称模式。在Be-78中,LDH-A在NT20中降低并在治疗后进一步下降,而在VL-10中,NT20和T20均呈现显著增加,表明该虫株存在更严重的组织损伤和代谢功能障碍。因此,尽管LDH-A为两种感染所共有,但其调节在VL-10中远为显著,强化了其更具侵袭性的表型。
总体而言,大多数共有蛋白在至少一种虫株的NT20组中上调,治疗效应在Be-78中更为明显,其中BNZ(T20)持续降低多种补体组分、免疫球蛋白和炎症标志物的丰度。在VL-10中,尽管治疗后一些标志物有所降低,但许多标志物仍远高于对照水平,反映了该虫株较低的治疗反应性。
在慢性期,70种蛋白在Be-78和VL-10的所有实验条件(CN、NT20和T20)中被共享(图6;表S10)。该集合表明,即使在晚期,两种感染仍维持一个共同的持续性免疫应答、组织重塑和与疾病慢性化相关的代谢改变轴。
在共享的免疫组分中,若干补体系统成员突出,包括C1qA、C1qB、C1qC和C1s-1,以及C反应蛋白——一个经典的慢性炎症标志物。广泛的重链、κ链和λ链免疫球蛋白库在两种虫株中保持调节,表明慢性期持续体液激活;然而,调节幅度在VL-10中普遍更高,尤其是κ链和J链。
结构重塑和组织支持蛋白也在共享组分中广泛存在,包括肌营养不良蛋白、肌球蛋白XVIIIb、α-内收蛋白、着丝粒相关蛋白E、含EGF腓骨蛋白样蛋白1和细胞骨架角蛋白42。这些蛋白的存在和调节提示细胞骨架、细胞外基质和组织组织的长期改变,在感染VL-10虫株的动物中效应更为显著,其丰度水平常超过感染Be-78组。
代谢和细胞损伤标志物强化了虫株间的不对称性。LDH-A在VL-10 NT20组中呈现剧烈增加,并在治疗后保持升高,而在Be-78组中其丰度降低或接近对照水平。同样,与炎性小体激活相关的Pyrin蛋白在两种虫株中均升高,但在VL-10感染组中达到更高水平,提示该模型中更强烈的慢性炎症激活。
综上,慢性期数据显示,尽管Be-78和VL-10虫株感染小鼠血清中存在一组共有蛋白,但蛋白质组调节幅度在感染VL-10虫株的动物中显著更大,炎症、结构和代谢蛋白丰度高。BNZ治疗(T20)后,Be-78感染小鼠在大多数共有蛋白中呈现更显著的降低,接近对照组谱图,而VL-10感染组维持持续高水平,表明持续全身炎症、活跃组织重塑和治疗反应有限(图7)。
更多结果和补充图表:doi:10.3390/microorganisms14030588
长按二维码关注我们,用最短的时间和最高的效率学习更多数据分析方法!
扫描上方二维码或登录平台官网后添加CNSknowall客服微信咨询!官网地址:
https://cnsknowall.com
CNSknowall:24年最新问世的遥遥领先的科研数据(0代码生信+统计学)分析平台,同时含有机制图模块+汉化版Pubmed融合Deepseek高效筛选目标文献+SCI文献例句/语料检索模块+OPenAI官方GPT接口,>500款CNS级别图表皆可一秒内一键出图,登录即秒变数据分析大神,体验前所未有的便捷数据分析之旅,开启科研天骄之路!
可向下滑动批阅!