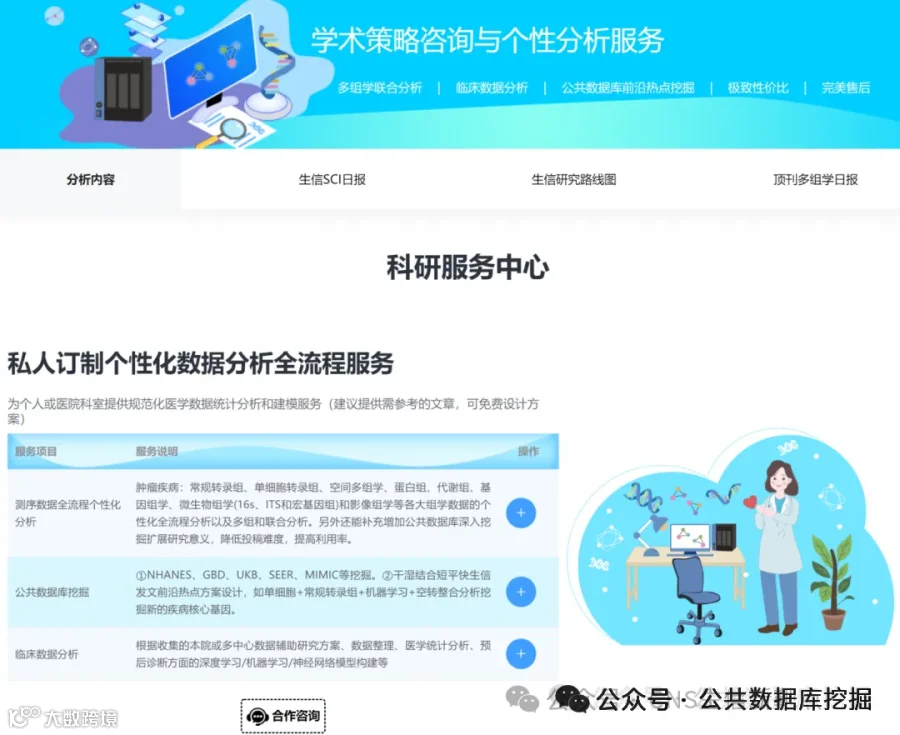
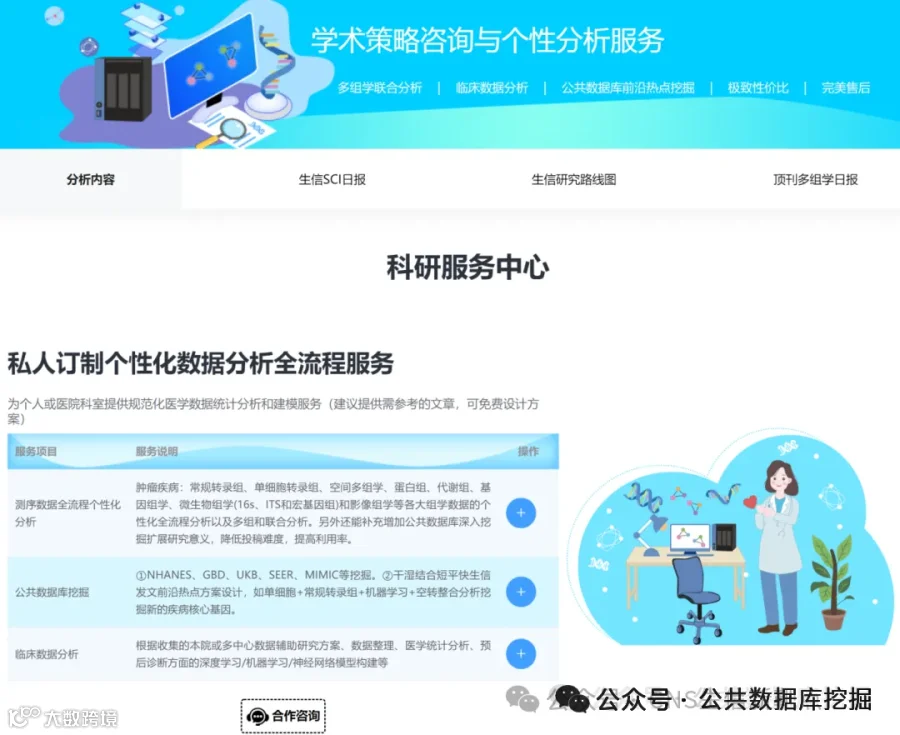
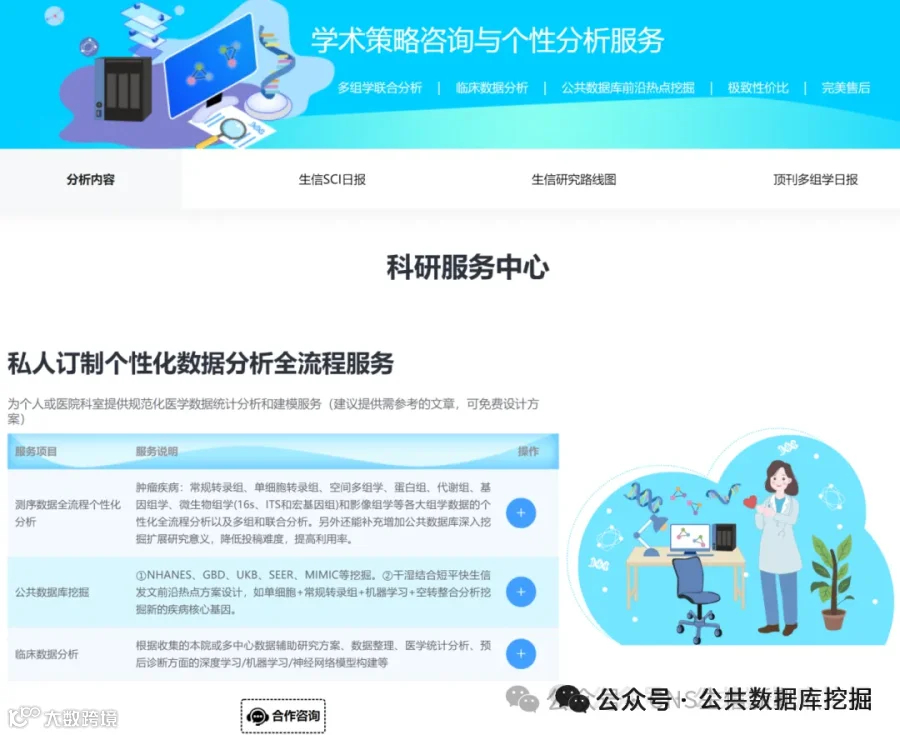
射血分数保留的心力衰竭(HFpEF)占所有心衰的一半以上,却一直缺乏有效治疗。其背后的罪魁祸首,可能就藏在心脏最微小的血管里!最新发表于《EMBO Molecular Medicine》的研究,通过单心脏内皮细胞转录组学、谱系示踪和AAV基因治疗,首次揭示:心脏微血管内皮细胞上的清道夫受体SR-B1是防止T细胞“攻击”心脏的关键屏障。在HFpEF中,SR-B1显著下降,导致转录因子IRF1激活,驱动内皮细胞大量分泌趋化因子CXCL10。CXCL10如同GPS信号,引导携带CXCR3受体的T细胞大量浸润心肌,引发舒张功能障碍和纤维化。研究团队在公共数据库GEO中验证了这一发现,并在HFpEF患者血浆中证实CXCL10水平与疾病严重度正相关(AUC诊断价值优异)。更令人振奋的是,通过AAV1病毒载体向内皮细胞回补SR-B1,可以完全逆转心脏病理重塑!这项研究为HFpEF提供了“SR-B1-CXCL10-CXCR3”全新治疗靶点。
今天给大家解读一篇4月发表在《EMBO Molecular Medicine》上的题目为“Microvascular endothelial scavenger receptor class B type I protects against heart failure with preserved ejection fraction by inhibiting T-cell cardiotropism.”的文章。本文研究心脏微血管内皮细胞(CMECs)功能障碍在射血分数保留的心力衰竭(HFpEF)中的机制。发现清道夫受体SR-B1在CMECs中高表达,并在HFpEF中显著下调。内皮特异性SR-B1缺失导致HFpEF小鼠模型中心脏病理性重构和舒张功能障碍加重,而通过AAV1重建SR-B1可缓解。机制上,SR-B1缺失通过激活IRF1信号增加CXCL10分泌,招募CXCR3阳性T细胞至心脏,从而恶化HFpEF。人类心脏组织及血浆分析证实该轴在临床样本中被激活。(请持续关注我们,每天为您解读最新见刊的文献!)想薅生信资料羊毛?直接在对话框回复 “资料”,免费领取干货大礼包!包括数据集、绘图代码、图表复现、思路总结、参考文献……0代码!鼠标点点点即可轻松完成5-10分生信SCI全文复现!
不想做实验,没数据,还想要快速发表文章,没问题的!公共数据库就是我们的数据宝藏!没思路不用担心,作为专业的生信团队,我们很乐意为你们效劳,提供研究路线设计和数据挖掘分析,扫码联系我们吧!



团队成员合影(位于上海陆家嘴中心,可随时预约参观)
题目:《微血管内皮清道夫受体B型I通过抑制T细胞心肌趋向性来预防射血分数保留的心力衰竭》Microvascular endothelial scavenger receptor class B type I protects against heart failure with preserved ejection fraction by inhibiting T-cell cardiotropism
发表期刊:EMBO Molecular Medicine
影响因子:8.3
研究背景:
心脏微血管内皮细胞(CMECs)功能障碍是射血分数保留的心力衰竭(HFpEF)的公认介质,但其潜在机制尚不清楚。已有研究提示清道夫受体在血管功能中发挥调节作用,但SR-B1在HFpEF中的具体角色未知。
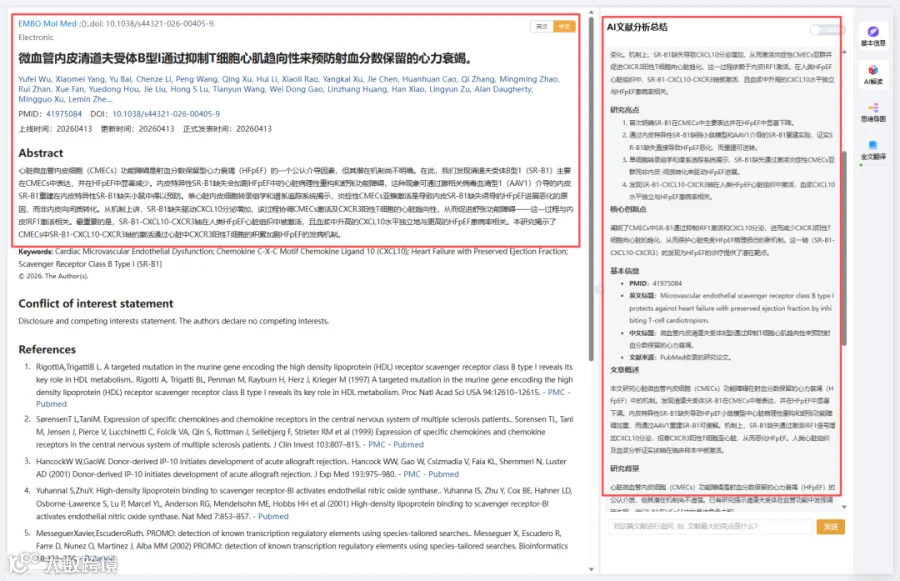
CNSknowall 平台 Pubmed+AI 快速提炼全文要点
研究思路:
-
检测SR-B1在HFpEF小鼠模型和正常小鼠CMECs中的表达水平。
-
利用内皮特异性SR-B1基因敲除小鼠(SR-B1-ECKO)构建HFpEF模型,观察心脏病理和舒张功能变化。
-
通过AAV1介导的SR-B1重建实验验证其保护作用。
-
采用单心脏内皮细胞转录组学和谱系追踪系统分析CMECs亚群变化。
-
体外及体内实验探索SR-B1缺失后下游信号(CXCL10、IRF1、CXCR3+T细胞)的变化。
-
在人类HFpEF心脏组织和血浆中验证SR-B1-CXCL10-CXCR3轴的激活及临床相关性。
研究亮点:
-
首次明确SR-B1在CMECs中主要表达并在HFpEF中显著下降。
-
通过内皮特异性SR-B1缺陷小鼠模型和AAV1介导的SR-B1重建实验,证实SR-B1缺失直接导致HFpEF恶化,而重建可逆转。
-
单细胞转录组学和谱系追踪系统揭示,SR-B1缺失通过激活炎症性CMECs亚群而非内皮-间质转化来驱动HFpEF进展。
-
发现SR-B1-CXCL10-CXCR3轴在人类HFpEF心脏组织中激活,血浆CXCL10水平独立与HFpEF患病率相关。
研究结果:
-
SR-B1主要在CMECs中表达,在HFpEF小鼠中显著降低。
-
内皮特异性SR-B1缺失加剧HFpEF的心脏病理性重构和舒张功能障碍,而AAV1介导的SR-B1重建可预防上述变化。
-
单细胞转录组学显示,SR-B1缺失导致炎症性CMECs亚簇激活,而非内皮-间质转化。
-
机制上,SR-B1缺失驱动CXCL10分泌增加,激活IRF1,进而促进CXCR3阳性T细胞向心脏趋化,导致舒张功能障碍。
-
人类HFpEF心脏组织中SR-B1-CXCL10-CXCR3轴被激活,血浆CXCL10水平升高且独立与更高的HFpEF患病率相关。
研究总结:
研究表明,CMECs中SR-B1的缺失通过激活IRF1-CXCL10轴,招募CXCR3阳性T细胞至心脏,从而加剧HFpEF的发病进程。SR-B1-CXCL10-CXCR3轴在人类HFpEF中同样被激活,提示该通路可能成为HFpEF诊断和治疗的新靶点。研究还指出,血浆CXCL10水平可作为HFpEF的独立生物标志物。这些发现深化了对HFpEF微血管功能障碍机制的理解,并为开发基于内皮保护的干预策略提供了理论基础。
结果译文:
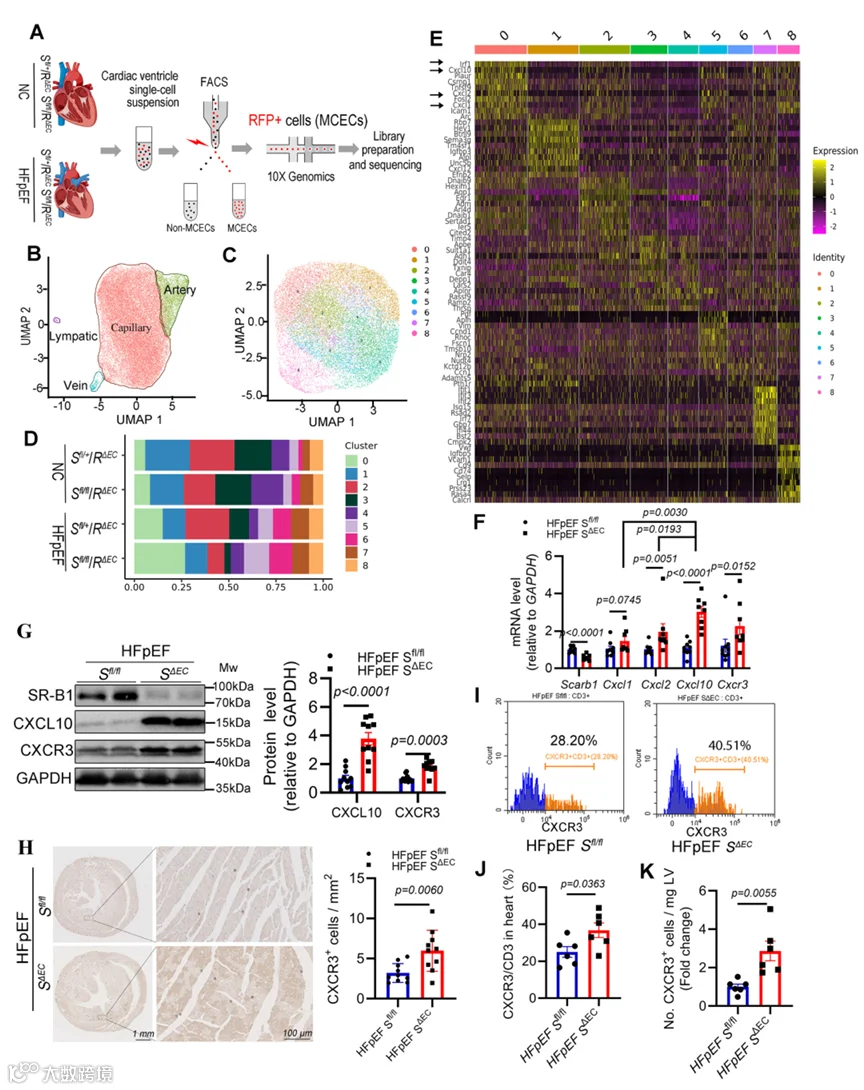
1.心脏微血管内皮SR-B1参与HFpEF发病机制
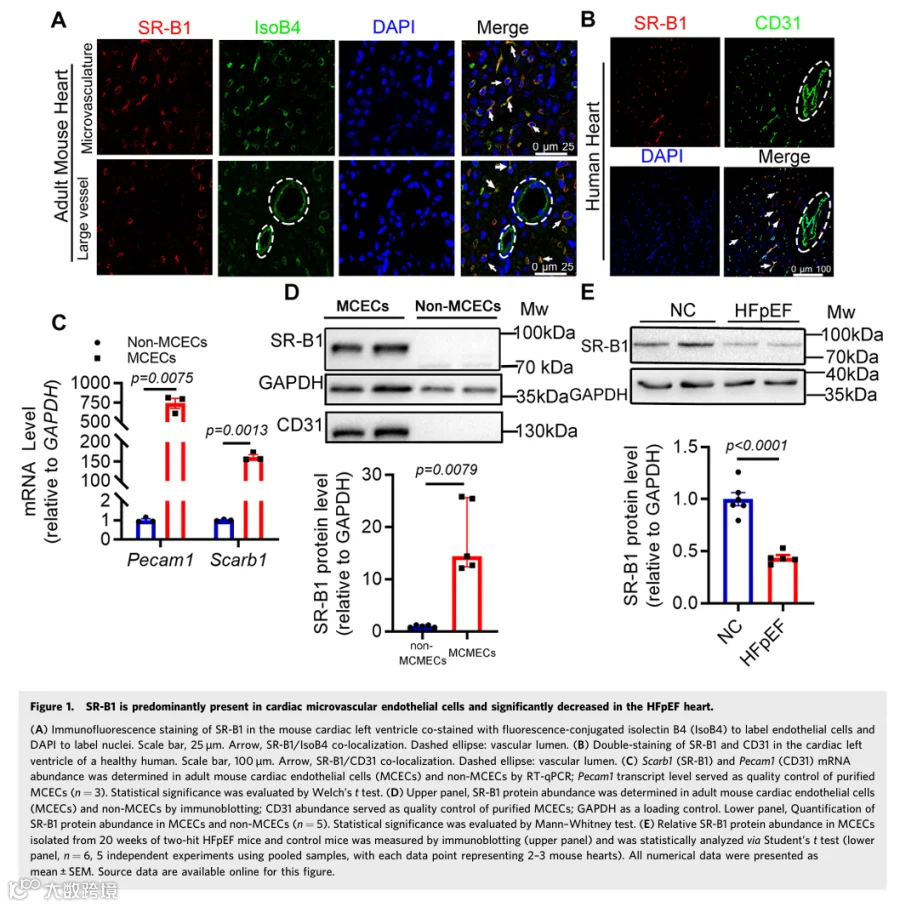
免疫荧光分析揭示SR-B1主要存在于小鼠和人心室心脏微血管内皮细胞(CMECs)中(图1A,B;附录图S1A,B)。我们验证了SR-B1在从小鼠心室分离的CD31阳性内皮细胞中高度丰沛(图1C,D;附录图S1C)。SCARB1(SR-B1)中的单核苷酸多态性rs10846742与心力衰竭发病机制显著相关(附录图S1D),表明其在体内可能参与HFpEF和HFrEF。随后,在HFpEF或HFrEF发病过程中评估了内皮SR-B1表达。首先,在体外用HFpEF样刺激处理的原代小鼠心脏内皮细胞中,SR-B1表达显著降低(附录图S1E)。一致且重要的是,与对照相比,从"两次打击"HFpEF小鼠分离的心脏内皮细胞中SR-B1蛋白丰度显著降低(图1E)。然而,由TAC诱导的HFrEF小鼠心脏内皮细胞中SR-B1转录保持不变(附录图S1F)。重要的是,小鼠心脏内皮细胞中Scarb1 mRNA表达水平依次降低,与HFpEF相关心功能障碍的恶化平行(附录图S1G)。因此,心脏CMECs中的SR-B1可能参与HFpEF发病机制。
2.内皮特异性SR-B1缺乏加重HFpEF中的舒张功能障碍
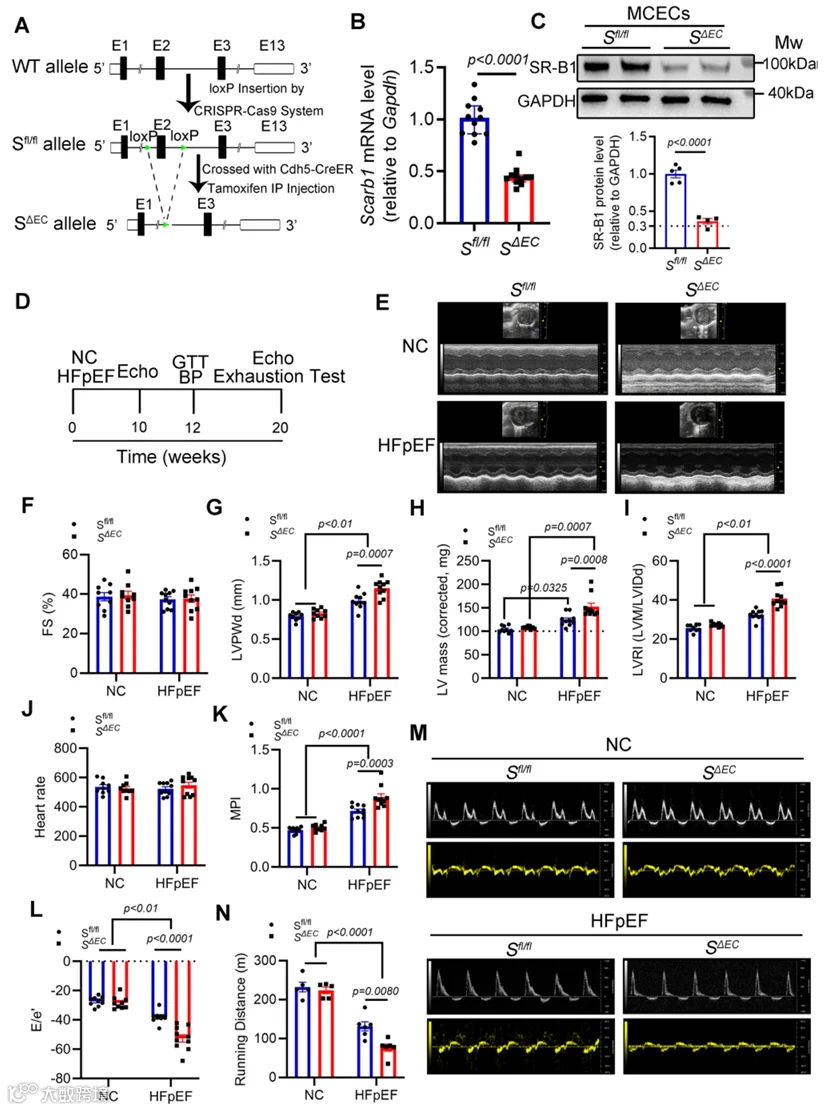
为直接研究内皮SR-B1在HFpEF发病机制中的作用,基于他莫昔芬诱导型Cre-loxP系统构建了内皮特异性SR-B1敲除小鼠(S0/0;Cdh5 CreER: SACE)(图2A)。简言之,利用CRISPR-Cas9基因编辑技术在小鼠Scarb1(SR-B1)基因第2外显子(158 bp)的侧翼内含子中插入loxP序列,构建了SR-B1flox/+(S0/+)小鼠。随后,将他莫昔芬诱导型内皮特异性钙粘蛋白5(Cdh5)-CreER小鼠与S0/+小鼠杂交,产生S0/+ Cdh5-CreER小鼠。然后,从S0/+ Cdh5-CreER小鼠与S0/+小鼠的杂交中获得S0/+和同窝对照S0/0小鼠,并通过DNA基因分型确认(附录图S2A)。与先前报道相反,此loxP插入策略未破坏SR-B1的内源性表达(附录图S2B)。最后,评估了SR-B1丰度和敲除特异性,证明内皮SR-B1被特异性删除,他莫昔芬注射后2周观察到心脏内皮细胞中约70%的降低(图2B,C;附录图S2C,D)。
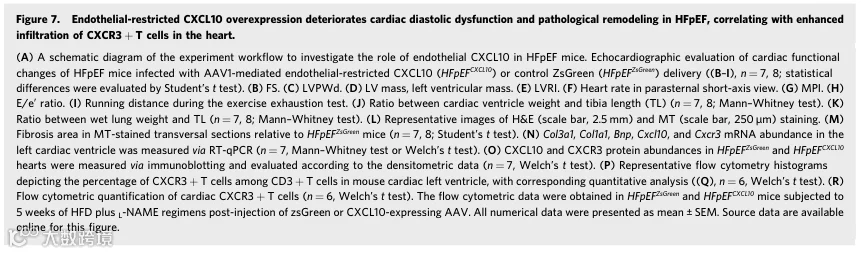
随后,我们研究了内皮SR-B1在HFpEF小鼠(即S0/0 HFpEF和S0/0 HFpEF小鼠)中的作用。经过10周HFD加L-NAME方案后,与对照S0/0和S0/0小鼠相比,S0/0 HFpEF和S0/0 HFpEF小鼠均表现出显著的左心室肥厚和舒张功能障碍,伴有正常的缩短分数(FS),确认了我们HFpEF模型的成功建立(附录图S3A-G)。值得注意的是,与S0/0 HFpEF小鼠相比,S0/0 HFpEF小鼠表现出趋向于更严重的心脏肥厚(通过LV质量和左心室重塑指数(LVRI)参数证明)和更差的舒张功能障碍(通过心肌性能指数(MPI)和E/e'比值指示),但这些差异未达到统计学显著性。
显著地,在将HFD和L-NAME方案延长至20周后,SABC HFpEF小鼠表现出比SII/I HFpEF小鼠显著加重的心脏肥厚和舒张功能,表明在缺乏内皮SR-B1的情况下疾病进展更快。具体而言,超声心动图显示显著增加的左心室舒张期后壁厚度(LVPWd)、LV质量、LVRI、MPI和E/e'比值,且FS正常(图2D-M)。此外,SABC HFpEF小鼠在应激运动测试中跑动距离显著缩短(图2N)。数据表明内皮SR-B1缺失加速了不良HFpEF重塑和进展。
3.内皮特异性SR-B1缺失恶化HFpEF中的心脏病理重塑,而SR-B1恢复可减轻功能障碍并防止SR-B1缺陷小鼠的不良重塑
目前研究表明,全局SR-B1缺乏能强力增加血浆HDL胆固醇(HDL-C)和总胆固醇(TC)浓度,并在高LDL胆固醇浓度下引起心功能障碍。然而,我们观察到内皮特异性SR-B1缺失不影响SABC与同窝对照S0/0小鼠的血浆HDL-C、TC和甘油三酯(TG)浓度(附录图S3H),与近期报道一致。此外,来自NC或HFpEF组的SABC和S0/0小鼠表现出相似的体重、血压(高血压)和葡萄糖耐量能力(肥胖)(附录图S3I-M)。数据表明SABC小鼠中最差的心脏舒张功能障碍不太可能是由于全身底物代谢(胆固醇、脂质和葡萄糖)和血压所致。
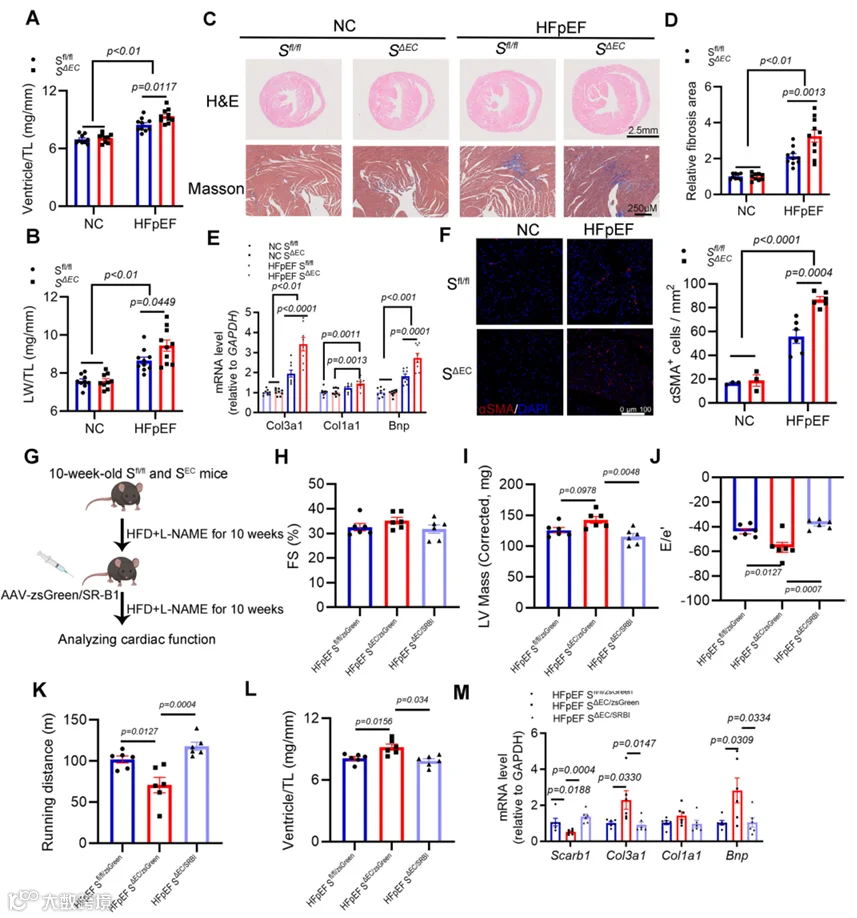
为进一步证明内皮特异性SR-B1缺乏对HFpEF心脏结构和功能的影响,在20周饮食干预后进行了心脏组织学分析和免疫组织学分析。结果显示,与S0/0小鼠相比,SABC HFpEF小鼠的心室重量与胫骨长度之比(Ventricle/TL)和肺湿重与胫骨长度之比(LW/TL)显著增加(图3A,B)。此外,与同窝对照相比,SABC HFpEF小鼠心脏纤维化程度显著增加(图3C-F)。因此,内皮SR-B1缺失促进了HFpEF中的心脏病理重塑。
为确定观察到的表型是否可通过在心脏内皮细胞中恢复SR-B1来挽救,我们采用了腺相关病毒1(AAV1)介导的内皮递送系统。基于先前文献,我们预选了四种AAV血清型进行初步筛选:AAV1、AAV2、AAV9和突变体RGDIRVS-AAV9。为实现在内皮细胞中的特异性表达,我们利用Cdh5启动子(一种成熟的内皮细胞特异性标志物)(附录图S4A)。通过注射后3周的免疫荧光和Western blot分析,我们确定AAV1血清型结合Cdh5启动子构建体表现出最佳的转导效率和特异性,用于后续基因递送实验(附录图S4B-E)。
对SABC和同窝对照S0/0小鼠进行HFD加L-NAME处理10周,随后通过尾静脉注射AAV1-Cdh5表达zsGreen或SR-B1(图3G)。注射后10周,通过超声心动图和运动耐力测试进行全面的心功能评估。结果表明SR-B1恢复显著改善了SR-B1缺乏引起的加重心脏肥厚、舒张功能障碍和运动诱导的疲劳(图3H-K)。此外,并且尤为重要地,病理学检查和RT-qPCR分析确认SR-B1重建显著抑制了SR-B1缺陷小鼠中心脏肥厚和心脏纤维化的进展(图3L,M)。总之,这些发现共同证明AAV1介导的SR-B1恢复有效挽救了与内皮SR-B1缺乏相关的病理表型。
4.内皮SR-B1在由横主动脉缩窄(TAC)手术诱导的急性HFrEF发病机制中是可有可无的
为研究内皮SR-B1在HFrEF中的作用,对SABC和S0/0对照小鼠进行了TAC手术。使用超声血流多普勒模块测量了主动脉弓结扎部位的血压,验证了两个TAC组(S0/0 TAC和SABC TAC小鼠)在缩窄部位显示相同的跨缩窄压力梯度(附录图S5A)。手术后2周,两个TAC小鼠组均发展为严重的扩张性心力衰竭伴收缩功能障碍,表现为FS显著降低(平均FS < 20%,附录图S5C)和LVIDD增加(附录图S5F)。值得注意的是,超声心动图分析揭示,在此急性HFrEF模型中心内皮SR-B1缺乏并未改变心脏收缩功能障碍或重塑的进展(附录图S5B-I)。此外,病理学检查和组织学分析证明SR-B1敲除未显著影响TAC诱导的心脏肥厚(附录图S5K)、肺水肿(附录图S5L)或纤维化(附录图S5M和5N)。通过RT-qPCR进行的分子标志物分析(包括Bnp、Col3a1和Col1a1)确认两个HFrEF组之间纤维化或肥厚标志物无显著差异(附录图S5O)。
为明确评估SR-B1的时间性影响,我们对一批新的内皮特异性SR-B1 KO小鼠进行了TAC。术后第2、4、6和8周的超声心动图揭示各基因型之间心功能无显著差异(附录图S5P-S)。生存分析显示小鼠从术后第2周开始死亡,至第8周死亡率高达30%(附录图S5T)。考虑到这一增加的死亡风险,小鼠被安乐死。病理学检查揭示SR-B1缺乏未显著改变心脏肥厚或肺水肿(附录图S5U,V)。总之,内皮SR-B1缺乏似乎不影响由TAC诱导的急性HFrEF的疾病进展。
5.谱系追踪系统显示,内皮向间充质转化在体内由内皮SR-B1缺失加重的HFpEF心脏舒张功能障碍中不起作用
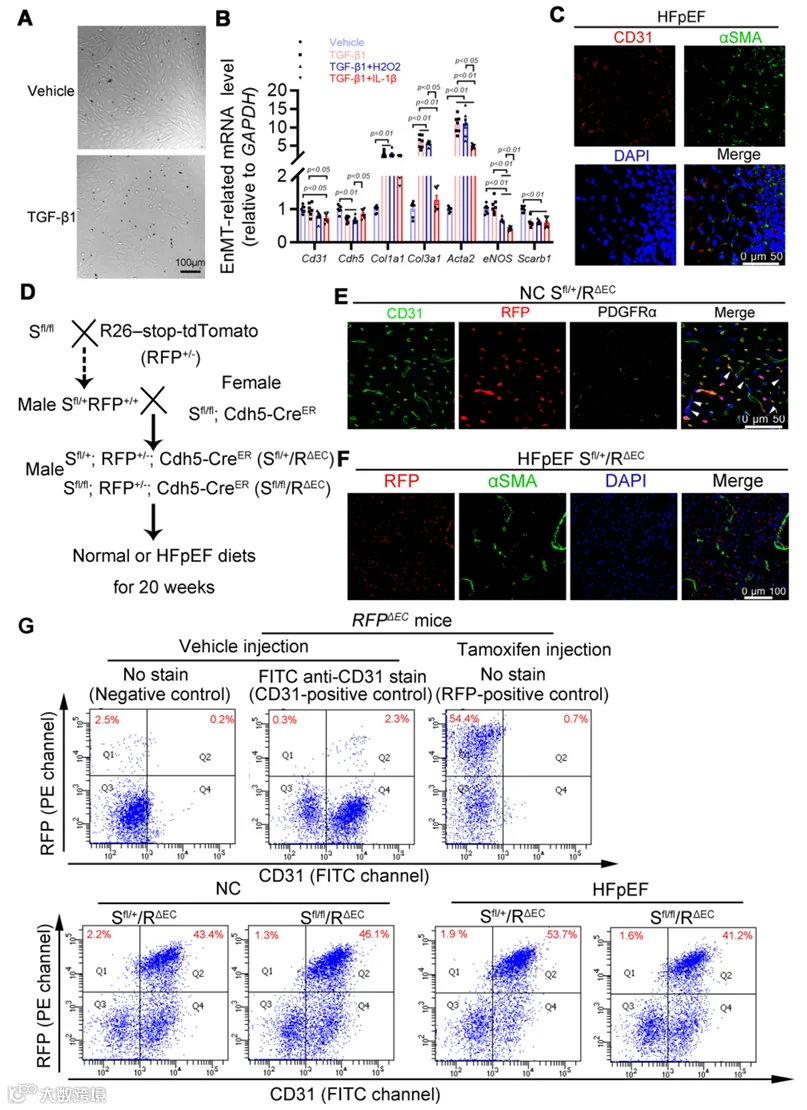
HFpEF的一个关键特征是全身性炎症,伴随高浓度的IL-1β、IL-6和肿瘤坏死因子-α(TNF-α)。由于体内内皮-间质转化(EnMT)也由类似的促炎因子驱动,我们有兴趣探究成年小鼠心脏内皮细胞是否具有EnMT潜力并参与HFpEF中的纤维化发展,更重要的是,这一过程是否由内皮SR-B1缺失所驱动。当用TGF-β1刺激从野生型小鼠分离的原代成年小鼠心脏内皮细胞时,其细胞形态发生了显著变化(图4A),伴随内皮标志基因Cdh5的显著下调和肌成纤维细胞主要特征基因的上调,如Col1a1、Col3a1和Acta2(αSMA)(图4B)。当与促炎因子H₂O₂或IL-1β及TGF-β1共孵育时,成年心脏内皮细胞表现出内皮标志物(包括CD31和内皮型一氧化氮合酶(eNOS))表达的显著降低(图4B)。值得注意的是,与单独孵育TGF-β1相比,IL-1β(而非H₂O₂)抑制了特征性肌成纤维细胞标志物(包括Acta2和Col3a1)的转录上调。此外,TGF-β1处理(无论是单独还是与H₂O₂或IL-1β联合)均显著降低了Scarb1(SR-B1)的转录。这些体外发现提示成年心脏内皮细胞具有EnMT的潜力,且SR-B1可能参与这一过程。
随后,我们在体内检验了HFpEF中的EnMT。在心脏纤维化区域使用双标记免疫荧光检测,未发现同时表达内皮标志物CD31和肌成纤维细胞标志物αSMA的细胞(图4C)。此外,我们构建了三重转基因谱系示踪小鼠模型,以可视化追踪HFpEF中心脏内皮细胞的潜在EnMT命运,并探究SR-B1是否参与EnMT过程(图4D)。在Sfl/fl/RΔEC小鼠和对照Sfl/+/RΔEC小鼠中,心脏内皮细胞在他莫昔芬注射后将表现出红色荧光蛋白(RFP)的持续特异性表达,同时伴有内皮特异性SR-B1敲除。随后,免疫荧光分析揭示在健康对照小鼠(NC Sfl/+/RΔEC)中,他莫昔芬注射后2周,RFP表达仅定位于心脏CD31阳性内皮细胞,而不存在于血小板源性生长因子受体α(PDGFRα)阳性的心脏成纤维细胞中(图4E)。值得注意的是,在进行20周HFD加L-NAME方案处理的谱系示踪Sfl/+/RΔEC小鼠中,心脏组织中未观察到αSMA与RFP阳性细胞的共定位(图4F)。接下来,使用荧光激活细胞分选(FACS)进一步确认HFpEF心脏中不存在EnMT。与免疫荧光共定位数据一致,在两个HFpEF组心脏中均检测不到RFP阳性但CD31阴性的细胞(图4G;附录图S6A)。因此,即使在缺乏SR-B1的情况下,EnMT也并非HFpEF小鼠心脏纤维化发展的原因。
6.内皮SR-B1缺乏导致炎症性CMEC亚簇增加伴CXCL10高表达
为研究内皮SR-B1缺乏加重HFpEF中不良心脏重塑和舒张功能障碍的分子机制,进行了scecRNA-seq以表征从三重转基因小鼠纯化的内皮细胞的转录谱(图5A)。首先,根据先前建立的单细胞转录组图谱,心脏内皮细胞被分类为毛细血管内皮细胞、动脉内皮细胞、静脉内皮细胞和淋巴内皮细胞(图5B;附录图S7A)。四个内皮细胞亚簇前10位标志物的热图分析揭示了不同的特征(附录图S7B)。接下来,我们对来自健康对照小鼠(NC S0/0/R0/0)各心脏内皮细胞亚簇中Scarb1(SR-B1)的表达进行了定量分析。结果表明Scarb1主要且在心脏毛细血管内皮细胞中显著表达(附录图S7C,D),这与组织学观察结果一致。此外,我们分析了四组小鼠毛细血管内皮细胞中Scarb1的表达(附录图S7E)。结果表明与单侧floxed对照相比,双侧floxed小鼠中Scarb1表达降低(NC S0/0/R0/0 vs NC S0/0/R0/0或HFpEF S0/0/R0/0 vs HFpEF S0/0/R0/0),确认了Scarb1的成功敲除。
由于SR-B1在CMEC中的主导表达,选择毛细血管内皮细胞簇进行进一步评估。层次(无监督)聚类鉴定了九个亚簇,投射到二维均匀流形逼近与投影(UMAP)图上(图5C)。组成分析揭示,毛细血管亚簇0的比例在HFpEF组中增加,由内皮SR-B1缺失驱动(图5D);亚簇3表现出相反的效应(图5D)。富集的京都基因与基因组百科全书(KEGG)通路热图显示,亚簇0中的特征标志基因与促炎通路相关,包括TNF-、IL-17-和NF-kappa B-信号通路,而这些通路在亚簇3中相对不活跃(附录图S7F)。亚簇0中前几位差异表达基因主要包括趋化因子,如Cxcl10、Cxcl2和Cxcl1(图5E)。此外,亚簇0中激活的前三条差异通路与这些趋化因子的高表达密切相关(附录图S7G)。因此,我们提出SR-B1缺失通过直接上调关键趋化因子浓度来驱动炎症性CMEC亚簇的激活。
为检验这一假设,我们定量了HFpEF小鼠心脏组织中Cxcl10、Cxcl1和Cxcl2的转录水平。分析揭示,与Cxcl1和Cxcl2相比,Cxcl10是具有最高倍数变化的差异表达基因(图5F)。类似地,与S0/0 HFpEF心脏相比,SABC HFpEF心脏中CXCL10蛋白显著上调(图5G)。此外,CXCR3(CXCL10的特异性受体)的mRNA和蛋白丰度在SABC HFpEF心脏中也显著高于S0/0 HFpEF心脏(图5F,G)。
接下来,我们的免疫荧光染色结果确认CXCR3主要在心脏T细胞中表达(附录图S8A),与单细胞数据集一致(附录图S8B)。此外,免疫组织化学染色揭示,与S0/0 HFpEF对应组相比,SABC HFpEF小鼠左心室中CXCR3阳性细胞显著积累(图5H)。为进一步确认SR-B1缺乏促进心脏中CXCR3+ T细胞浸润,进行了流式细胞术分析(圈门策略见附录图S6B)。结果证明SR-B1缺乏增加了CXCR3+ T细胞/CD3+ T细胞比率(图5I,J),并增强了HFpEF心脏左心室中CXCR3+ T细胞(图5K)和总CD3+ T细胞(附录图S8C)的浸润,而不影响CD45+和CD11b+细胞群(附录图S8C)。
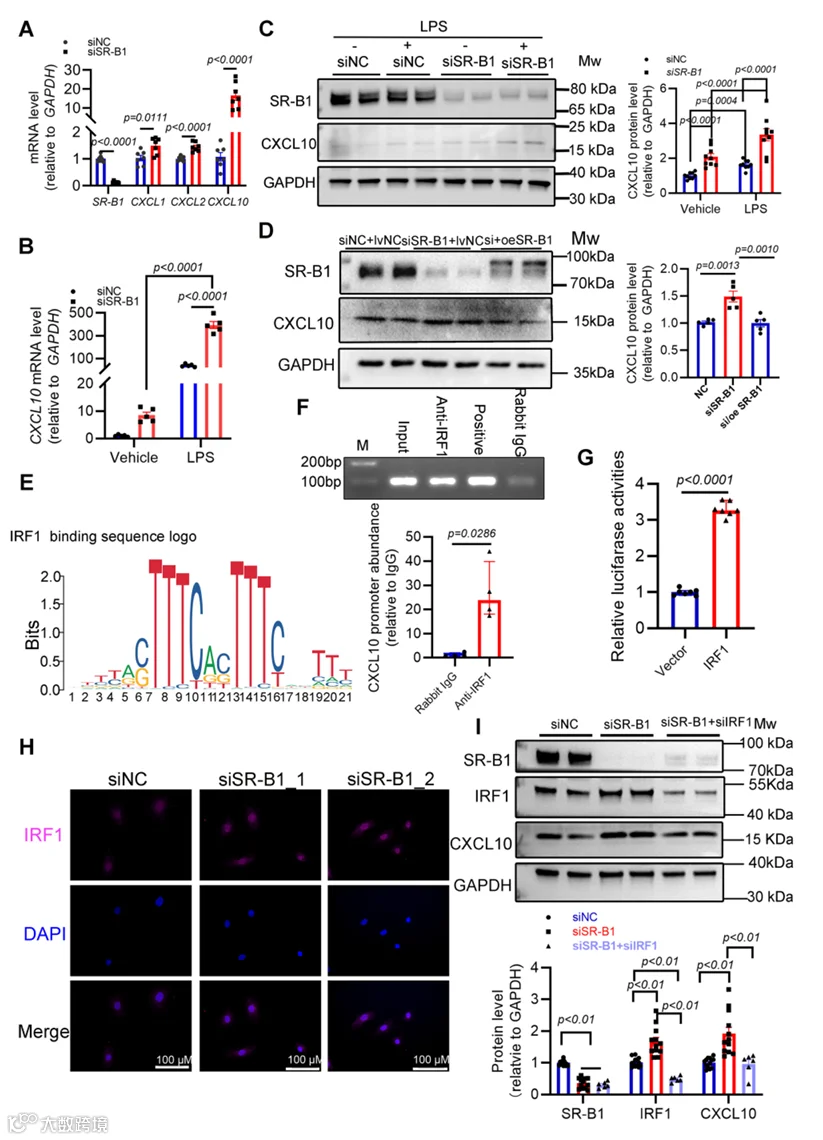
7.IRF1的激活可能是由SR-B1缺失引起CXCL10上调的原因
我们试图在原代人CMECs(HCMECs)中连接SR-B1和CXCL10。在SR-B1缺乏的HCMECs中,CXCL10转录以上调幅度大于CXCL1和CXCL2的倍数变化上调(图6A)。先前已有报道,作为HFpEF关键合并症的肥胖,可引起全身慢性炎症,具有显著高水平的內毒素。为确定反向SR-B1/CXCL10关系背后的机制,使用內毒素脂多糖(LPS)诱导HCMECs的炎症激活,当用LPS处理时CXCL10也在SR-B1敲低的HCMECs中显著上调(图6B,C)。此外,在HCMECs中重建SR-B1降低了CXCL10蛋白丰度(图6D)。发现表明SR-B1的存在阻止CXCL10表达,从而抑制炎症激活。
我们接下来使用PROMO工具v2.0探索了SR-B1介导的CXCL10调控的分子机制,预测结合CXCL10启动子的潜在转录因子。通过Ensemble网站鉴定的启动子序列(附录图S9A)被上传到PROMO进行分析,结果见附录图S9B。值得注意的是,IRF1在预测的转录因子中排名第九,但作为毛细血管内皮细胞促炎亚簇0中排名首位的标志基因出现(图5E)。随后,染色质免疫沉淀后qPCR(ChIP-qPCR)确认IRF1能够结合CXCL10启动子(图6E,F)。双荧光素酶实验进一步证明与对照相比,在IRF1过表达细胞中CXCL10启动子驱动的信号显著增加(图6G)。此外,免疫荧光染色表明IRF1在HCMECs中呈主要的细胞核定位,且在SR-B1缺乏时信号增强(图6H)。因此,我们假设IRF1直接介导SR-B1缺乏条件下CXCL10的转录上调。体外挽救实验验证了IRF1是SR-B1缺失后CXCL10蛋白上调所必需的(图6I)。
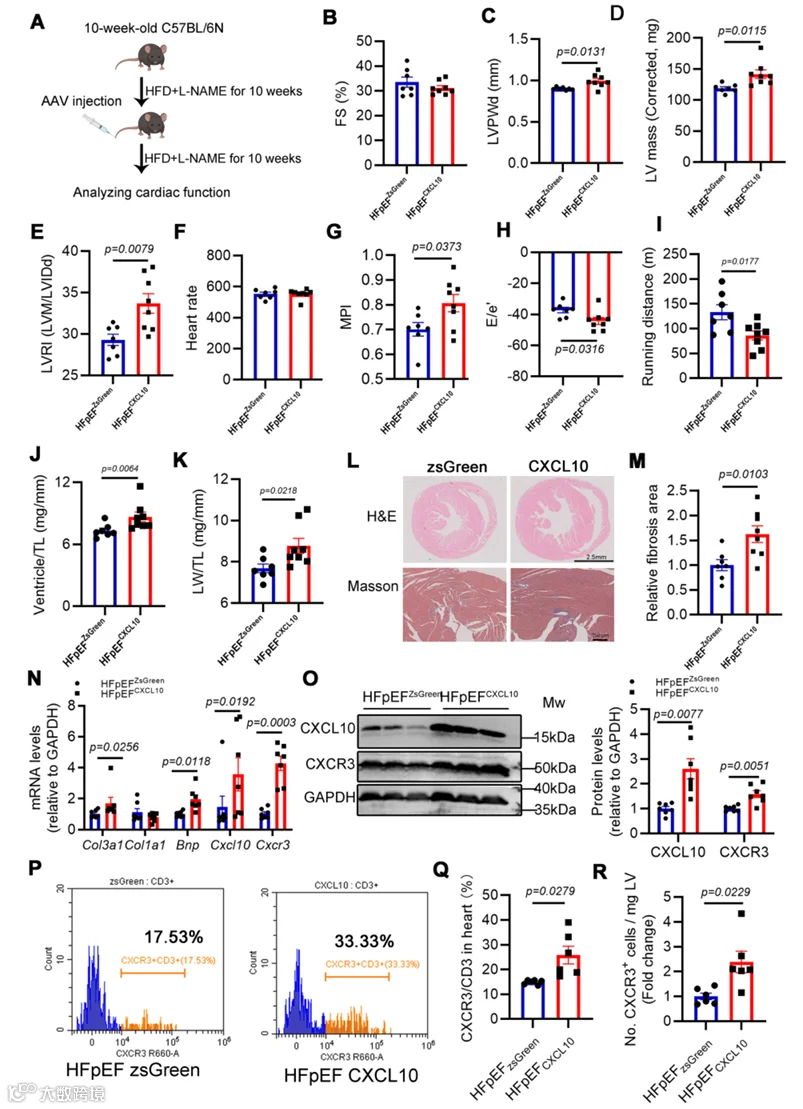
8.体内内皮限制性CXCL10过表达促进CXCR3+ T细胞向心脏浸润,并加重HFpEF中的心脏舒张功能障碍和病理重塑
我们试图通过内皮特异性AAV1递送研究CXCL10在SR-B1缺乏介导的HFpEF不良心脏重塑中的体内作用,并确定CXCL10是否足以将CXCR3+ T细胞募集到心脏组织中。对野生型小鼠进行HFD加L-NAME处理10周,之后通过尾静脉注射表达zsGreen或CXCL10的AAV1载体,再继续相同饮食方案10周(图7A)。超声心动图揭示两个HFpEF组之间FS保持正常(图7B)。然而,过表达内皮CXCL10的HFpEF小鼠(即注射AAV1-Cdh5-CXCL10的小鼠)比注射AAV1-ZsGreen的HFpEF小鼠表现出更严重的心肌重塑和舒张功能障碍(图7C–H)。运动力竭测试确认CXCL10过表达小鼠表现出显著缩短的跑动距离,表明心脏功能储备严重降低(图7I)。进一步的离体评估揭示,注射AAV1-CXCL10的HFpEF小鼠比注射AAV1-ZsGreen的HFpEF小鼠具有更严重的心室肥厚和肺水肿(图7J,K)。组织学和分子分析揭示,CXCL10过表达HFpEF小鼠的心脏表现出更严重的纤维化和肥厚(图7L–N)。因此,内皮选择性CXCL10过表达损害了心功能,这与SR-B1缺乏表型类似。
CXCL10在自身免疫性疾病和癌症中的病理生理作用已被充分阐明。由内皮细胞、成纤维细胞或巨噬细胞以旁分泌方式分泌的CXCL10,可将CXCR3+ T细胞募集到组织中,从而加剧自身免疫性炎症级联反应或抑制癌症进展。然而,尚不清楚是否特异性来源于心脏内皮细胞的CXCL10能够驱动CXCR3+细胞向心脏浸润。
首先,我们采用Western blot和RT-qPCR分析,发现与ZsGreen表达对照相比,CXCL10过表达心脏中CXCL10和CXCR3的表达显著升高(图7N,O)。接下来,我们利用流式细胞术在AAV注射后5周评估了HFpEF小鼠中的免疫细胞浸润(附录图S11A)。与AAV注射后10周的小鼠一致,这些CXCL10过表达小鼠表现出心脏肥厚和舒张功能障碍(附录图S11B–E)。流式细胞术结果证明,内皮CXCL10过表达导致CXCR3+/CD3+ T细胞比率和CXCR3+ T细胞计数显著增加(图7P–R),以及CD45+白细胞的显著升高(附录图S11F)。CD3+ T细胞和CD11b+髓系细胞保持不变(附录图S11G,H)。
随后,我们应用相同策略评估了CXCL10对健康小鼠免疫细胞浸润和心功能的影响(附录图S11I)。值得注意的是,这些小鼠未表现出显著的心脏肥厚或舒张功能障碍(附录图S11J–M)。流式细胞术分析揭示,除了CXCR3+/CD3+ T细胞比例的显著增加外,与注射AAV1-Cdh5-zsGreen的对照相比,注射AAV1-Cdh5-CXCL10的小鼠中免疫细胞亚群(包括CD45+、CD11b+、CD3+或总CXCR3+细胞计数)未观察到显著改变(附录图S11N–S)。
总之,CXCL10是HFpEF中致病性CXCR3+ T细胞浸润和心功能障碍的关键介质,但它不能独立在健康小鼠中诱导显著的免疫微环境变化或功能损伤。这提示CXCL10介导的CXCR3+ T细胞向心肌的募集可能依赖于全身性炎症信号与心脏特异性微环境因子之间的相互作用。
尽管女性在临床上对HFpEF的易感性更高,但在广泛使用的临床前两次打击模型中,雌性性别却矛盾性地具有保护作用。为探究这种保护是否延伸至微血管炎症激活,我们在去卵巢(OVX)的两次打击雌性小鼠中评估了关键分子(Irf1、Cxcl1、Cxcl2、Cxcl10、Icam1)。未检测到显著变化(附录图S12)。这种微血管激活的缺失与近期一篇报告一致,该报告显示XBP1s下调(在雄性小鼠中的一个关键发现)在雌性小鼠中不存在。因此,我们研究中缺乏分子变化可能确实归因于此特定模型中雌性小鼠的基础性保护机制。
9.临床HFpEF患者中IRF1-CXCL10/CXCR3轴与心脏舒张功能障碍密切相关
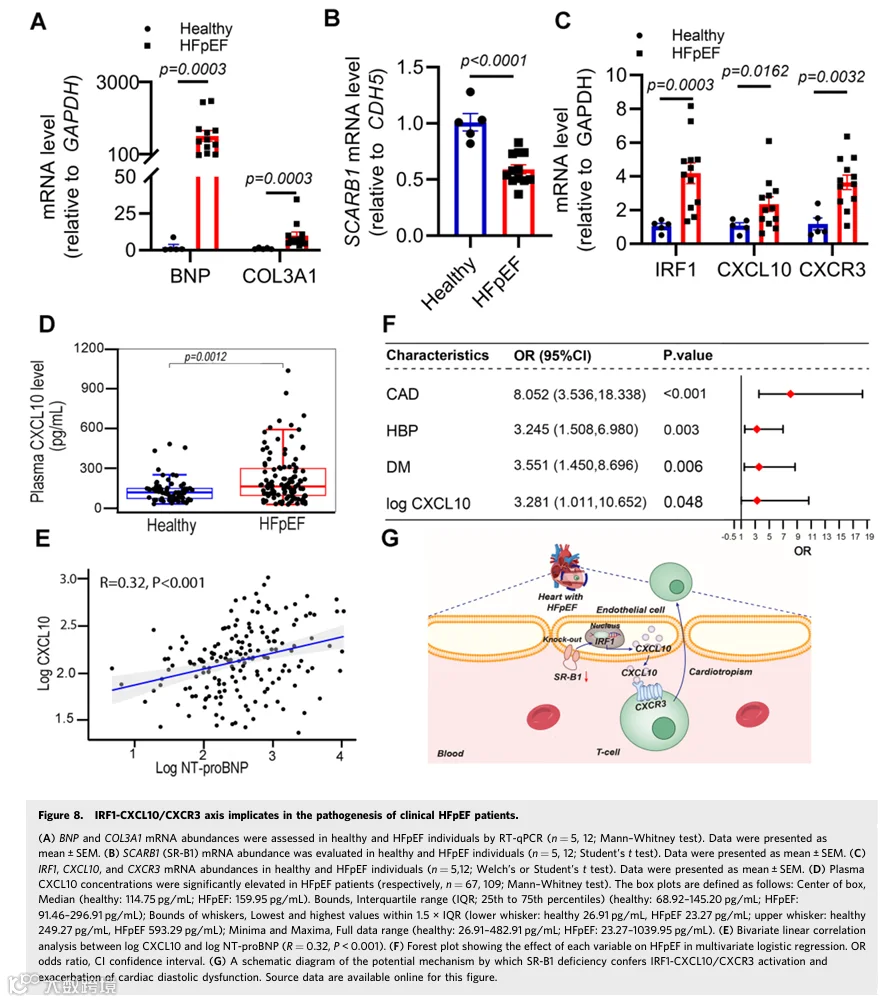
为支持这些临床前实验发现,在心肌病相关HFpEF患者的左心室中评估了SR-B1-IRF1-CXCL10/CXCR3轴的基因丰度。HFpEF供体的特征表明HFpEF患者具有保留的LVEF,但左心室舒张期后壁厚度(LVPWT)、E/e比值和NT-proBNP浓度增加(附录表S1)。实验数据表明,与健康对照组相比,HFpEF患者心室含有高丰度的BNP和COL3A1,与SRB1基因显著下调和IRF1、CXCL10和CXCR3丰度显著增加协同一致(图8A-C)。
我们还招募了176名个体(109例HFpEF病例和67例健康对照)来研究血浆CXCL10浓度与HFpEF之间的相关性(附录表S2)。使用酶联免疫吸附实验测量血浆CXCL10和N末端前B型利钠肽(NT-proBNP)浓度。与对照血浆相比,HFpEF血浆中CXCL10和NT-proBNP浓度显著增加(图8D;附录图S13A)。双变量线性相关分析揭示血浆CXCL10与NT-proBNP浓度之间存在正相关(r = 0.32, P < 0.001,图8E)。随后,进行单变量和多变量logistic回归分析以鉴定HFpEF的风险因素(附录表S3)。基于单变量和多变量logistic回归分析结果,log CXCL10(OR = 3.28, 95% CI: 1.01-10.65, P = 0.048)是与HFpEF正相关的因素(图8F)。此外,HFpEF血浆中HDL-C浓度显著较低(1.20 vs 1.01, P < 0.001;附录表S2)。额外的Spearman相关分析显示血浆CXCL10水平与HDL-C呈负相关(r = -0.18, P = 0.015;附录图S13B),提示HFpEF血浆中CXCL10丰度的增加可能与低水平HDL介导的SR-B1激活有关。发现表明IRF1-CXCL10/CXCR3轴参与临床HFpEF发病机制。
更多结果和补充图表:doi: 10.1038/s44321-026-00405-9
长按二维码关注我们,用最短的时间和最高的效率学习更多生信思路!
扫描上方二维码或登录平台官网后添加CNSknowall客服微信咨询!官网地址:https://cnsknowall.com
CNSknowall:24年最新问世的遥遥领先的颠覆性科研数据(0代码生信+统计学)分析平台,同时含有机制图模块(原创3000多素材和机制图模板)+AI一键生成高质量比国自然标书初稿+汉化版Pubmed融合Deepseek高效筛选目标文献同时一键提炼全文核心创新点+SCI文献例句/语料检索模块+全文翻译+文献求助+图片查重+期刊查询+OPenAI官方GPT接口,>500款CNS级别图表皆可一秒内一键出图,登录即秒变数据分析大神,体验前所未有的便捷数据分析之旅,开启科研天骄之路!
可向下滑动发掘更多科研秘籍!