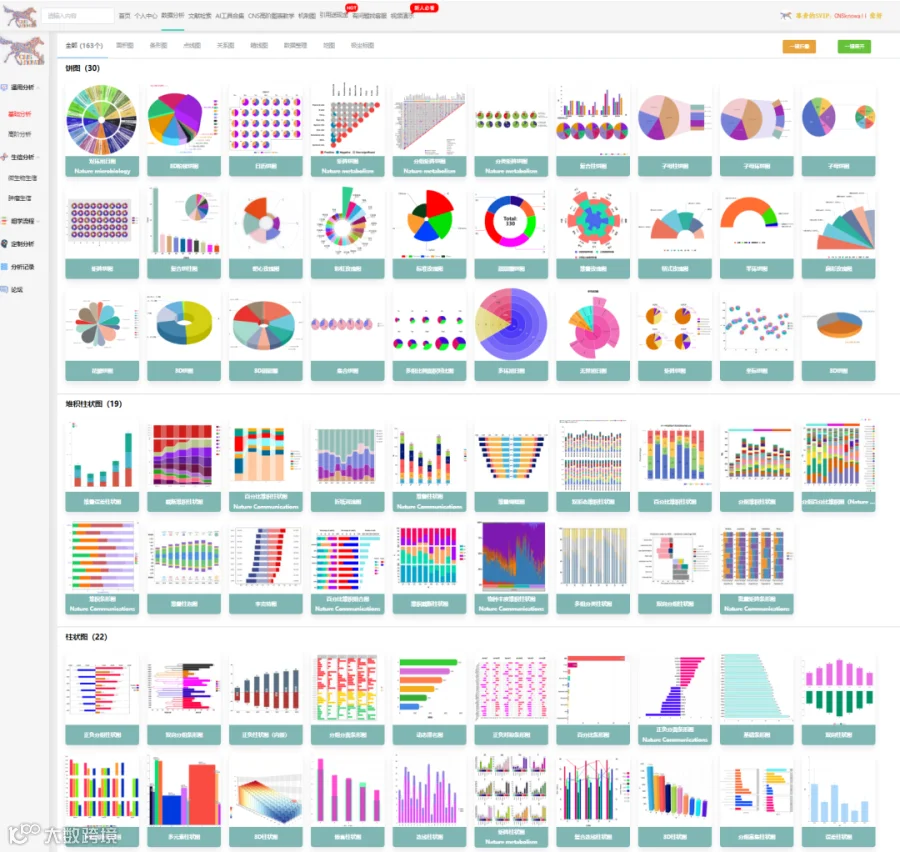
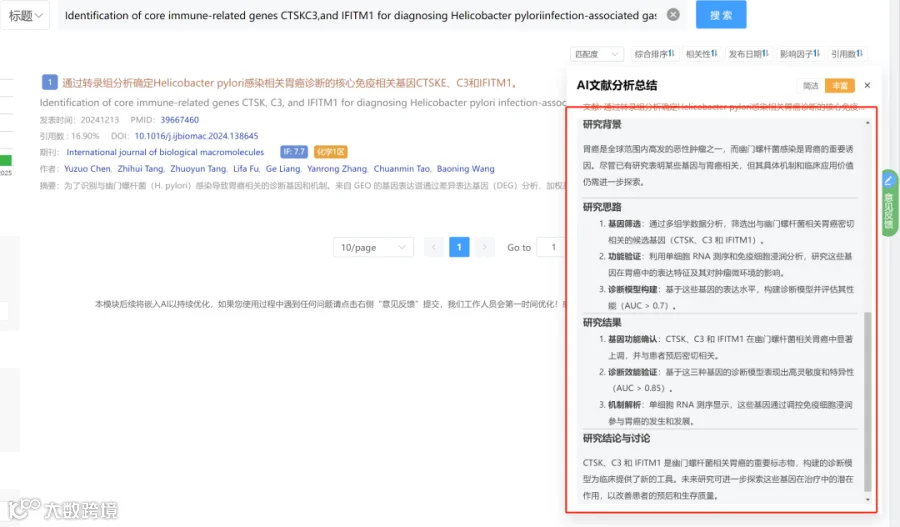
今天给大家解读一篇4月发表在《iScience》上的题目为“Transient tumor exposure induces persistent functional defects in memory CD8+ T cells.”的文章。本研究旨在探讨短暂的肿瘤暴露对记忆CD8+ T细胞功能的影响。研究人员使用表达相同抗原表位的牛痘病毒(VV-NP68)和肿瘤细胞(EL4-NP68)模型,分别模拟了急性病毒感染和肿瘤排斥过程。通过比较两种方式产生的记忆CD8+ T细胞(Vir-CD8+与Tum-CD8+),发现Tum-CD8+细胞在转录组和功能上均表现出显著缺陷。文献不仅描述了这些表型和功能差异,还通过二次应答实验证明了该缺陷的稳定性和不可逆性,从而强调了初始免疫启动环境对于塑造长期免疫记忆质量的关键作用。(请持续关注我们,每天为您解读最新见刊的文献!)想薅生信资料羊毛?直接在对话框回复 “资料”,免费领取干货大礼包!包括数据集、绘图代码、图表复现、思路总结、参考文献……0代码!鼠标点点点即可轻松完成5-10分生信SCI全文复现!
不想做实验,没数据,还想要快速发表文章,没问题的!公共数据库就是我们的数据宝藏!没思路不用担心,作为专业的生信团队,我们很乐意为你们效劳,提供研究路线设计和数据挖掘分析,扫码联系我们吧!









团队成员合影(位于上海陆家嘴中心,可随时预约参观)





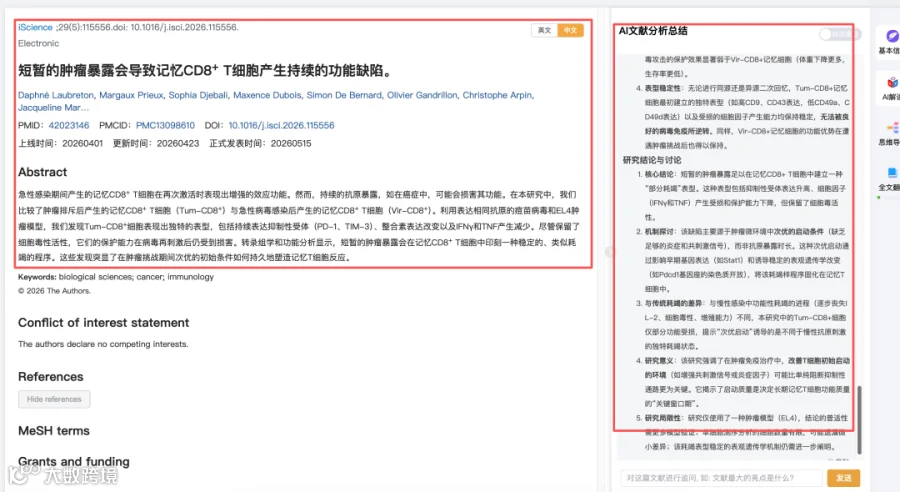
题目:《短暂的肿瘤暴露会导致记忆CD8+ T细胞产生持续的功能缺陷》Transient tumor exposure induces persistent functional defects in memory CD8+ T cells
发表期刊:iScience
影响因子:4.1
研究背景:
- 优良记忆的范例
急性感染后,大多数效应T细胞死亡,小部分分化为记忆T细胞。这些记忆细胞在再次遭遇抗原时能迅速增殖并执行强大的效应功能(如产生细胞因子和细胞毒性)。 - 不良记忆的后果
相反,在慢性感染或癌症等持续性抗原暴露条件下,CD8+ T细胞会进入一种称为“耗竭”的低应答状态,表现为功能受损和抑制性受体持续表达。 - 待解决的问题
肿瘤抗原通常缺乏适当的炎症和共刺激信号,T细胞启动效率低下。那么,在没有慢性抗原暴露的情况下,仅由无效共刺激信号导致的“次优启动”,是否足以影响记忆CD8+ T细胞的功能?这正是本研究要回答的问题。

研究思路:
- 模型建立
通过过继转移F5 TCR转基因T细胞,然后分别用表达NP68抗原的VV病毒(i.n.)或EL4肿瘤细胞(s.c.)免疫小鼠,确保两种挑战都能被清除。 - 表型与转录组对比
在记忆阶段,比较Vir-CD8+与Tum-CD8+细胞的表面标志物表达、单细胞转录组谱,寻找差异。 - 功能验证
在体外,比较两种记忆细胞的细胞因子(IFNγ, TNF, IL-2)产生能力、细胞毒活性和增殖能力。 - 体内保护功能评估
用流感病毒(Flu-NP68)进行二次攻击,比较两种记忆细胞对体重减轻和生存率的保护效果;并通过过继转移实验验证。 - 稳定性测试
给予二次同源或异源免疫(VV或EL4),评估初次挑战诱导的表型和功能特征是否会发生改变。
研究亮点:
- 明确缺陷来源
证明即使是短暂的、不持续的肿瘤抗原暴露,也足以在记忆CD8+ T细胞中烙印下一个不可逆的耗竭样程序。 - 揭示表型稳定性
这种耗竭样表型在肿瘤被清除后仍稳定存在,并且不能通过后续的优质免疫(如病毒感染)进行纠正。 - 与传统耗竭模型区分
该研究建立的这种由“短暂暴露”诱导的耗竭样程序,其特征(如保留细胞毒活性)与慢性感染导致的完全耗竭状态存在差异。
研究结果:
- 表型和转录组差异
Tum-CD8+细胞相较于Vir-CD8+细胞,表现出表达更低水平的整合素(CD49a, CD49d)及更高水平的抑制性受体(PD-1, TIM-3)和活化相关分子(CD43, CD9, Galectin-3)。单细胞转录组分析显示,两者基因表达模式相似,但Tum-CD8+细胞上调了与耗竭相关的基因(如Pdcd1, Havcr2),同时下调了与抗病毒、代谢和细胞粘附相关的基因(如Stat1, Itga4)。 - 功能缺陷
Tum-CD8+记忆细胞保留了细胞毒活性和IL-2产生能力,但其产生IFNγ和TNF的能力显著降低,且增殖能力受损。降低的细胞因子产量与肿瘤大小或抗原暴露时长无关。 - 保护能力受损
在随后的病毒感染中,Tum-CD8+记忆细胞向感染部位(如肺部组织)的迁移能力较差,导致其对致死性流感病毒攻击的保护效果显著弱于Vir-CD8+记忆细胞(体重下降更多,生存率更低)。 - 表型稳定性
无论进行同源还是异源二次回忆,Tum-CD8+记忆细胞最初建立的独特表型(如高CD9、CD43表达,低CD49a、CD49d表达)以及受损的细胞因子产生能力均保持稳定,无法被良好的病毒免疫所逆转。同样,Vir-CD8+记忆细胞的功能优势在遭遇肿瘤挑战后也得以保持。
研究总结:
- 核心结论
短暂的肿瘤暴露足以在记忆CD8+ T细胞中建立一种“部分耗竭”表型。这种表型包括抑制性受体表达升高、细胞因子(IFNγ和TNF)产生受损和保护能力下降,但保留了细胞毒活性。 - 机制探讨
该缺陷主要源于肿瘤微环境中次优的启动条件(缺乏足够的炎症和共刺激信号),而非抗原暴露时长。这种次优启动通过影响早期基因表达(如Stat1)和诱导稳定的表观遗传学改变(如Pdcd1基因座的染色质开放),将该耗竭样程序固化在记忆T细胞中。 - 与传统耗竭的差异
与慢性感染中功能性耗竭的进程(逐步丧失IL-2、细胞毒性、增殖能力)不同,本研究中的Tum-CD8+细胞仅部分功能受损,提示“次优启动”诱导的是不同于慢性抗原刺激的独特耗竭状态。 - 研究意义
该研究强调了在肿瘤免疫治疗中,改善T细胞初始启动的环境(如增强共刺激信号或炎症因子)可能比单纯阻断抑制性通路更为关键。它揭示了启动质量是决定长期记忆T细胞功能质量的“关键窗口期”。 - 研究局限性
研究仅使用了一种肿瘤模型(EL4),结论的普适性需更多模型验证;单细胞测序分析的细胞数量有限,可能遗漏微小差异;该耗竭表型稳定的表观遗传学机制仍需进一步阐明。
结果译文:
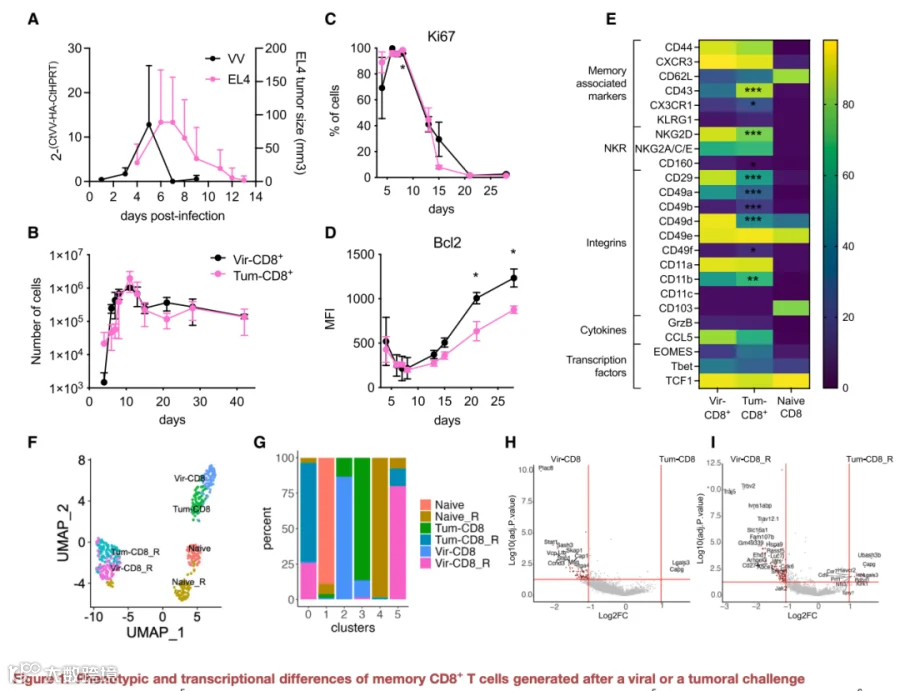
1.肿瘤或病毒诱导的记忆细胞展现出有限数量的表型和转录谱差异
2.肿瘤诱导记忆CD8+ T细胞表达与T细胞耗竭相关的分子
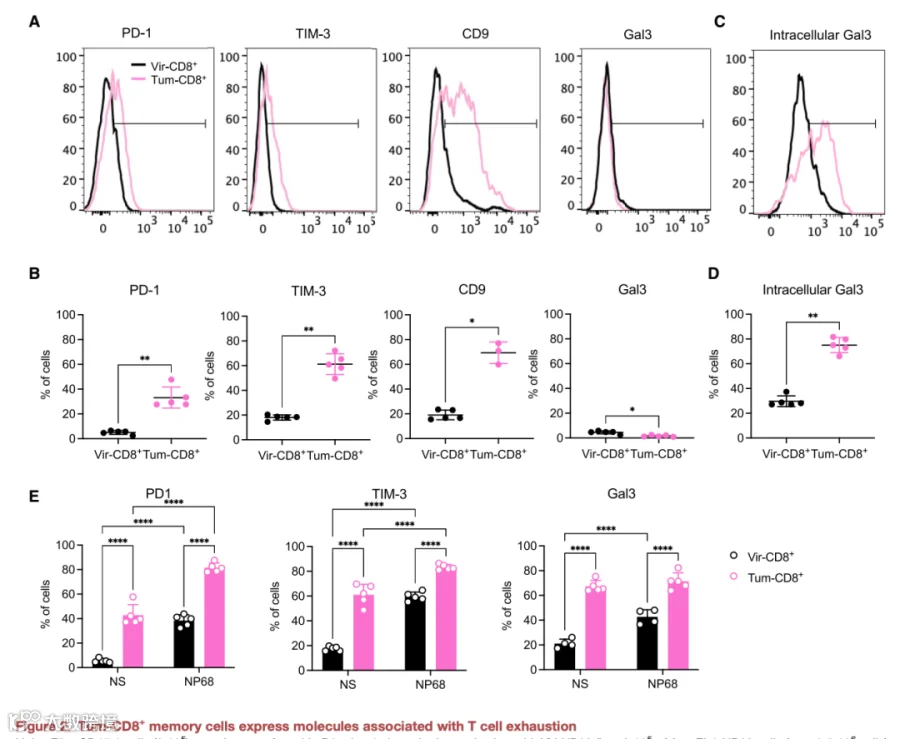
我们证实了静止或激活的肿瘤诱导记忆细胞过表达如Pdcd1(PD-1)、Havcr2(TIM-3)、Lgals3(Gal3)和Cd9(CD9)等基因。除Gal3外,所有这些标志物在静止肿瘤诱导记忆细胞中的表面蛋白表达增加均已通过验证,与病毒诱导记忆细胞相比(图2A及2B)。
3.肿瘤诱导记忆CD8+ T细胞的细胞因子产生能力受损,但细胞毒性得以保留
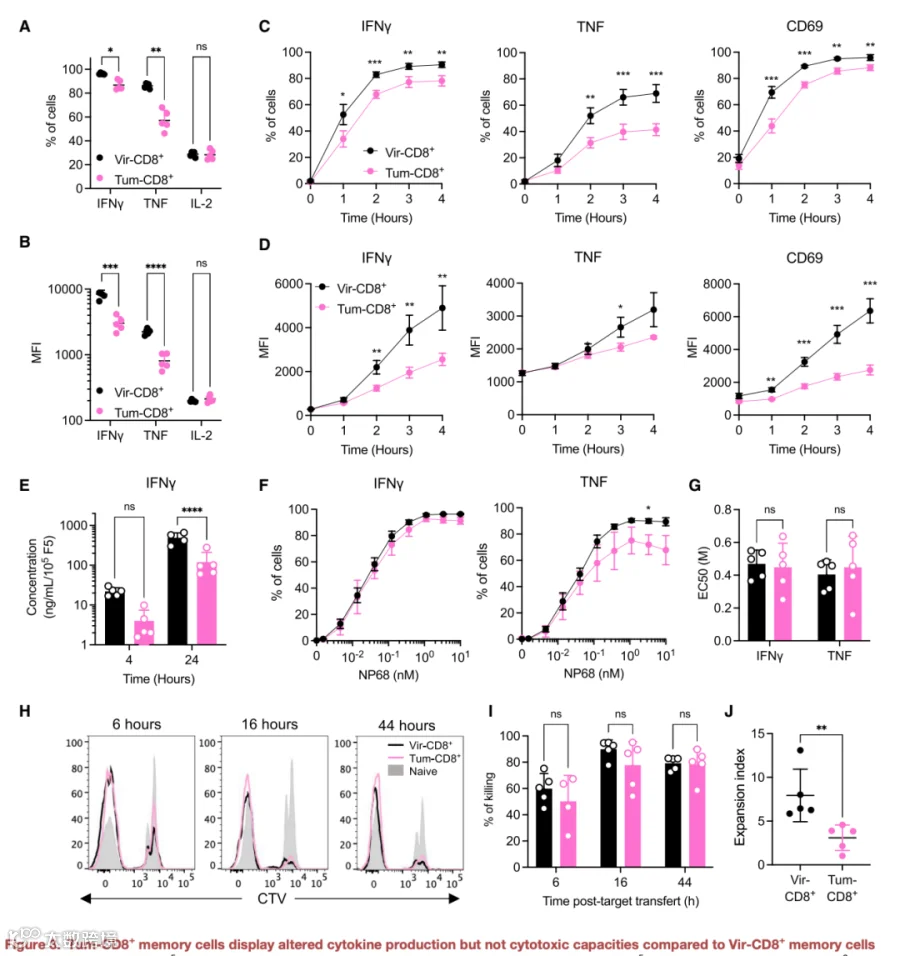
为了明确与肿瘤诱导记忆细胞中Gal3和抑制性受体表达差异相关的功能性后果,我们分析了它们的细胞因子产生和细胞毒性能力。我们首先证明,在体外用NP68刺激后,两种记忆细胞类型展现出相当的IL-2产生能力(图3A及3B)。相反,与病毒诱导记忆细胞相比,产生IFNγ和TNF的肿瘤诱导记忆细胞比例更小(图3A)。此外,基于平均荧光强度值,病毒诱导记忆细胞中的细胞内细胞因子水平显著更高(图3B),提示肿瘤诱导记忆细胞的细胞因子产生能力受损。这一点通过在细胞水平和上清液中进行的细胞因子产生时程分析得到进一步证实。事实上,与病毒诱导记忆细胞相比,肿瘤诱导记忆细胞中IFNγ和TNF的产生始终较低(图3C-3E)。此外,在所有时间点,肿瘤诱导记忆细胞表面表达的CD69水平均保持低于病毒诱导记忆细胞(图3C及3D)。重要的是,这种细胞因子产生的减少与抗原负荷或暴露时间无关(图S3E及S3I)。
4.短暂肿瘤挑战足以损害F5记忆细胞的保护能力
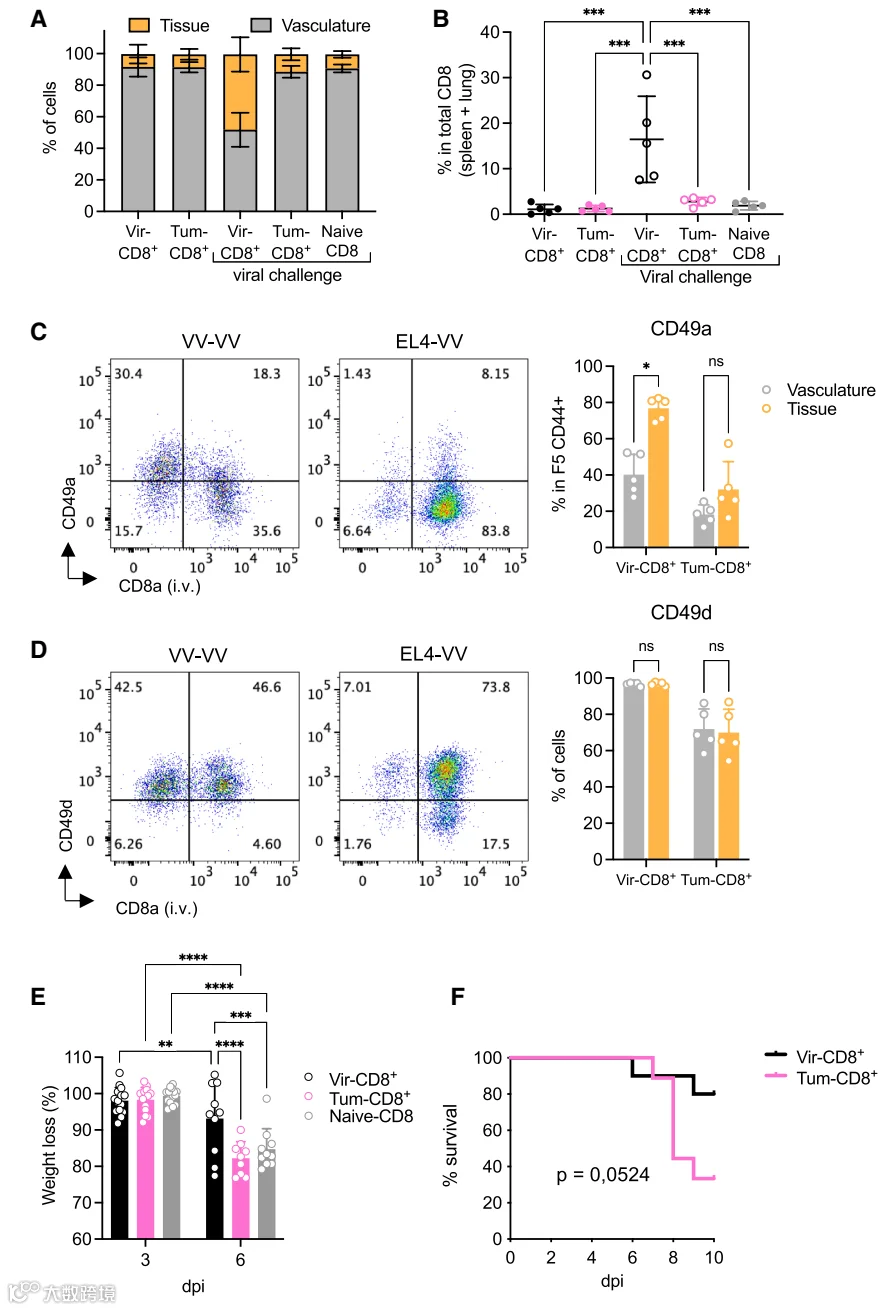
鉴于挑战期间病毒和肿瘤细胞均被清除,我们接下来测试了CD8+记忆细胞是否能够控制随后的病毒感染。在初次免疫后30天,用VV-NP68(鼻内)感染曾接受病毒或肿瘤挑战的小鼠。六天后,分析肺组织和脉管系统中F5 CD8+细胞的数目和表型。病毒挑战后,肺组织内肿瘤诱导记忆细胞的比例低于病毒诱导记忆细胞(图4A)。为评估记忆CD8+ T细胞被招募到肺的能力,我们确定了脾脏和肺中F5记忆细胞的总数(图S5),并计算了每只小鼠被招募至肺组织的这些细胞的比例。尽管稳态条件下肺组织内肿瘤诱导记忆细胞与病毒诱导记忆细胞的比例相似,但病毒挑战后,病毒诱导记忆细胞而非肿瘤诱导记忆细胞或幼稚细胞被强烈招募至肺组织(图4B)。这表明病毒诱导记忆细胞具有更好的迁移至感染部位的能力。这与肺内病毒诱导CD8+ T细胞(而非肿瘤诱导记忆细胞)上CD49a表达增加相关,CD49a是一种参与靶向细胞至肺的整合素(图4C)。然而,位于脉管系统和组织中的细胞之间,CD49d的表达并无差异(图4D)。

5.F5记忆细胞的表型和细胞因子产生能力在同源或异源回忆后得以保守
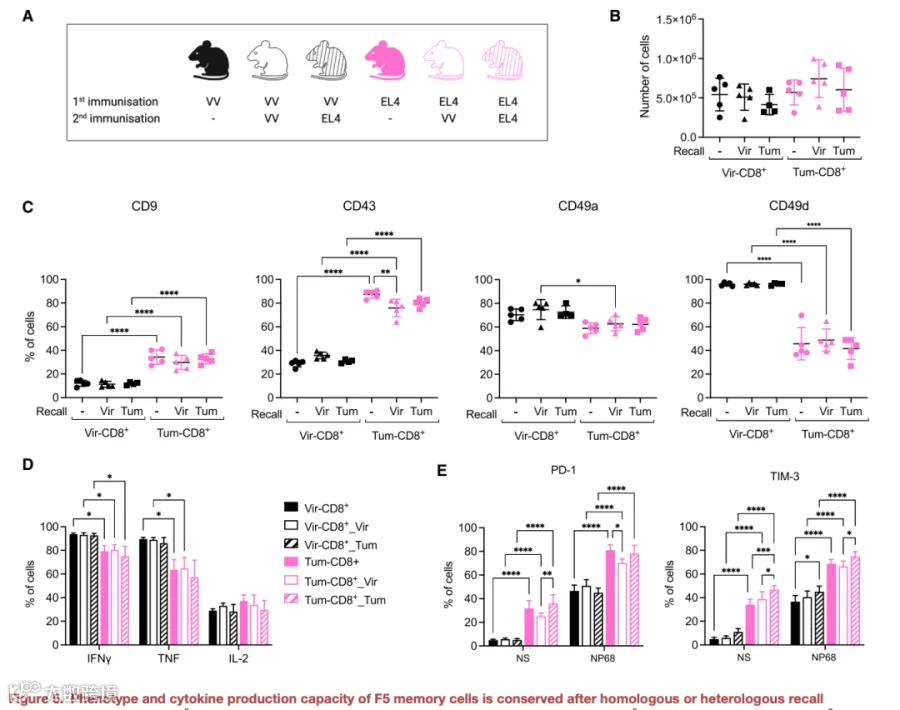
最后,我们试图评估病毒或肿瘤挑战后诱导的表型的稳定性。为此,在攻击后26天,对曾接受病毒或肿瘤挑战的小鼠进行第二次同源或异源回忆,使用VV-NP68或EL4-NP68(图5A)。未接受二次免疫的小鼠用作对照。在回忆后31天,从所有组别小鼠中回收了等量的F5记忆细胞(图5B)。在脾脏记忆细胞中测定了初次挑战后区分肿瘤诱导记忆细胞与病毒诱导记忆细胞的主要标志物的表达。病毒回忆后,病毒诱导记忆细胞中CD43的表达轻微增加;肿瘤回忆后,肿瘤诱导记忆细胞中CD43的表达无论使用VV-NP68还是EL4-NP68回忆均轻微降低(图5C)。然而,在所有回忆条件下,两组之间CD43表达的整体差异得以维持。同样,肿瘤诱导记忆细胞上PD-1的表达仍高于病毒诱导记忆细胞,且肿瘤回忆并未改变两种细胞类型间的差异(图5D)。此外,体外刺激后,肿瘤诱导记忆细胞产生IFNγ的水平低于病毒诱导记忆细胞(图5E)。这种缺陷不仅在同源(肿瘤+肿瘤)回忆后持续存在,在异源(病毒+肿瘤和肿瘤+病毒)回忆后也同样持续(图5E)。针对流感病毒的攻击,我们观察到两种细胞类型在回忆后保护存活的能力仍有显著差异(图5F及5G)。总之,在短暂肿瘤暴露后诱导的记忆CD8+ T细胞耗竭样表型被证实是稳定且不可逆的,即使是在随后由病毒挑战提供的强效TCR和炎症信号存在的情况下也是如此。
更多结果和补充图表:doi:10.1016/j.isci.2026.115556



扫描上方二维码或登录平台官网后添加CNSknowall客服微信咨询!官网地址:https://cnsknowall.com
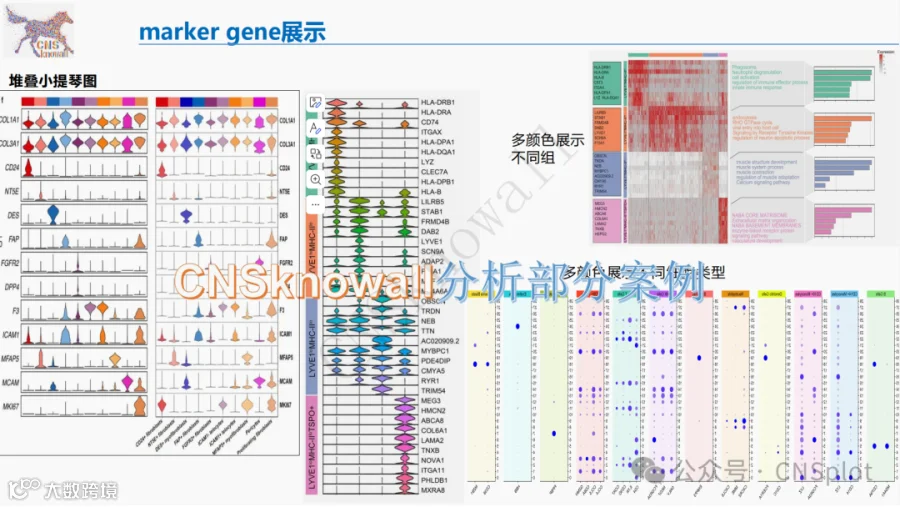
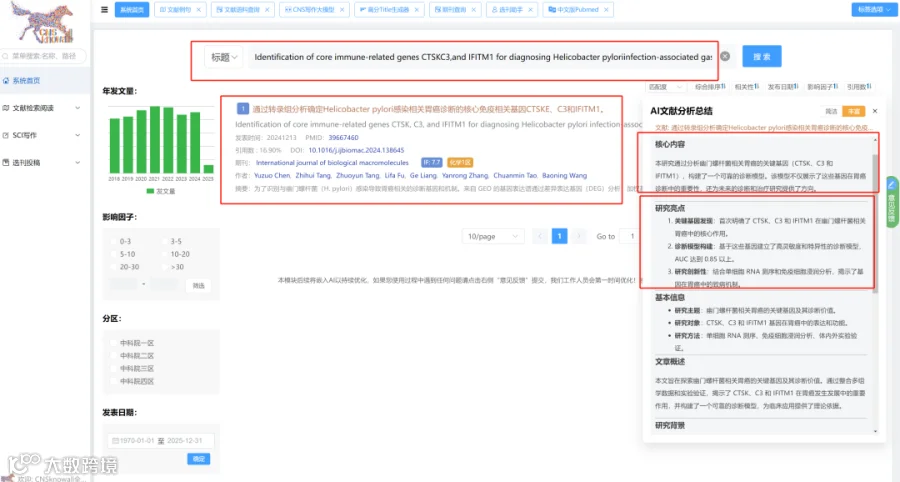
CNSknowall:24年最新问世的遥遥领先的颠覆性科研数据(0代码生信+统计学)分析平台,同时含有机制图模块(原创3000多素材和机制图模板)+AI一键生成高质量比国自然标书初稿+汉化版Pubmed融合Deepseek高效筛选目标文献同时一键提炼全文核心创新点+SCI文献例句/语料检索模块+全文翻译+文献求助+图片查重+期刊查询+OPenAI官方GPT接口,>500款CNS级别图表皆可一秒内一键出图,登录即秒变数据分析大神,体验前所未有的便捷数据分析之旅,开启科研天骄之路!
可向下滑动发掘更多科研秘籍!