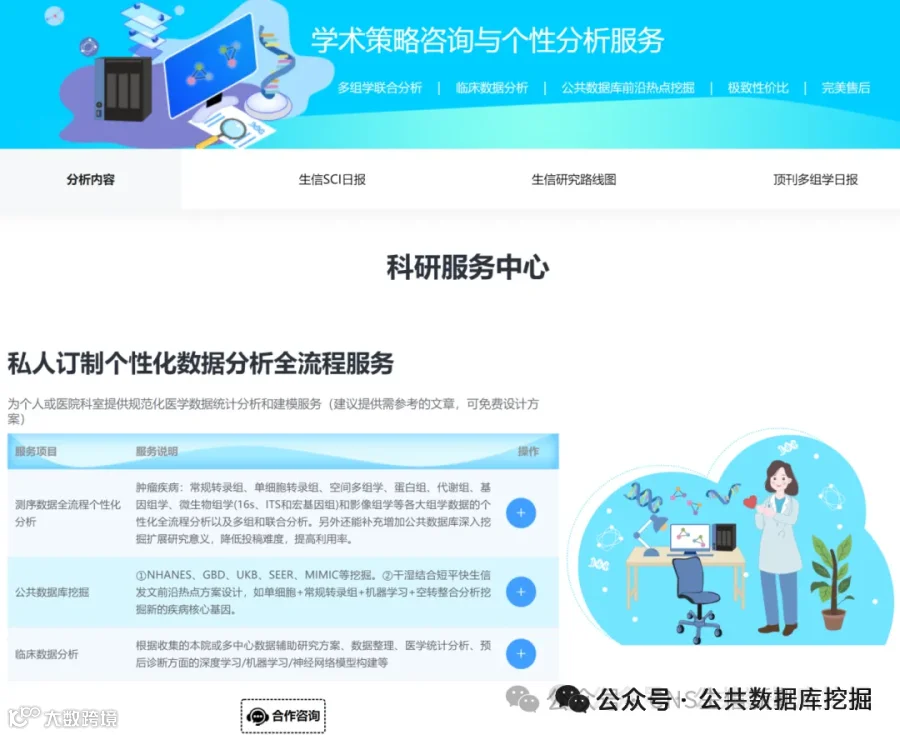
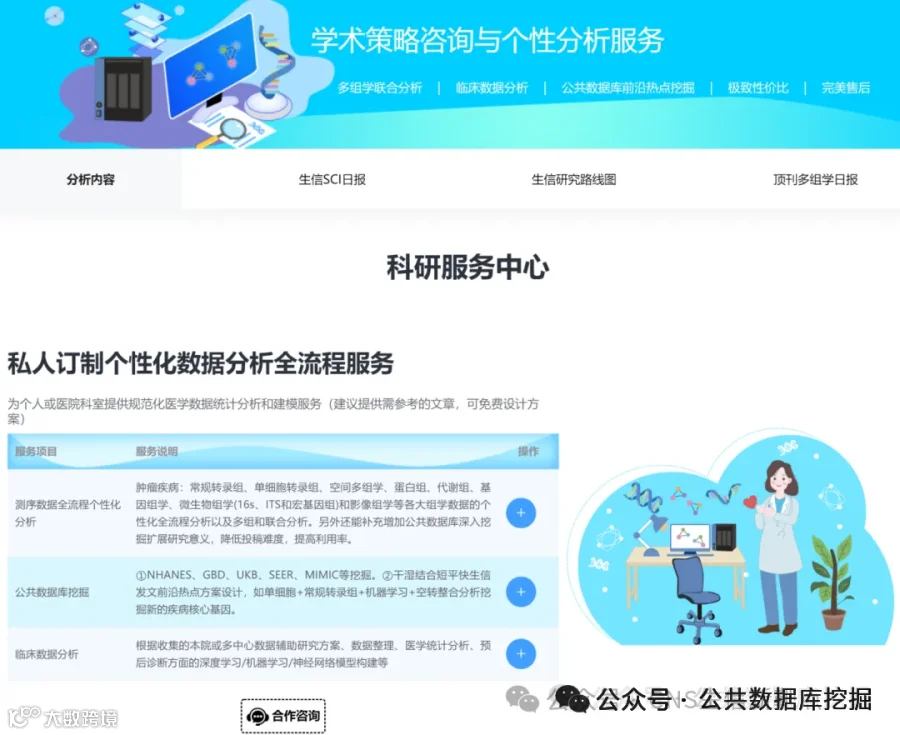
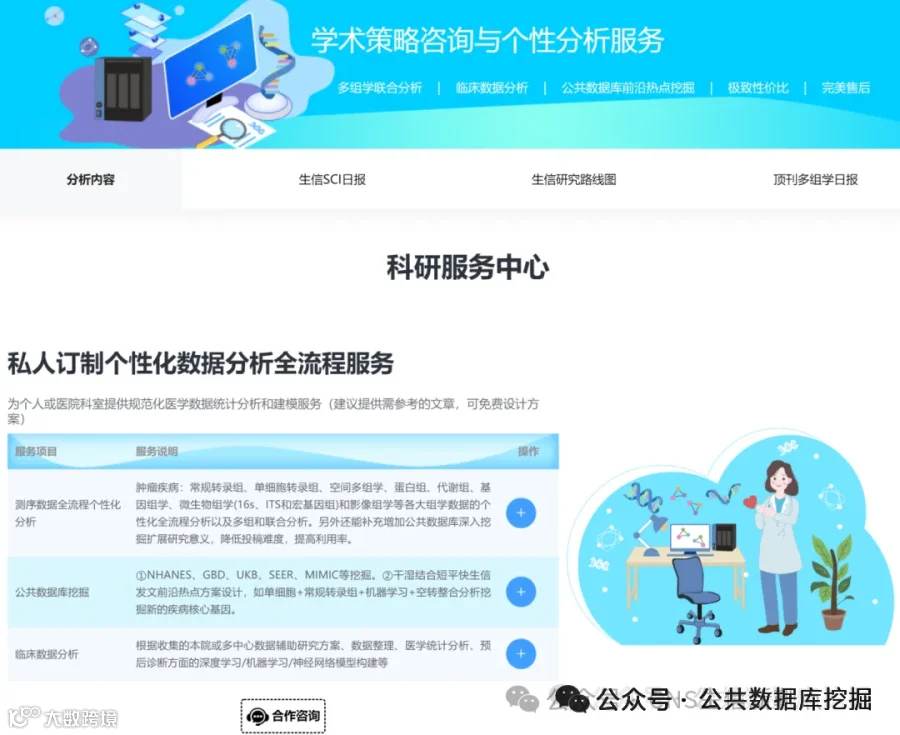
肛门直肠畸形(ARM)是新生儿常见的先天性畸形,全球发病率约1/5000,但其遗传基础尚不明确。本研究通过全外显子测序对8例综合征型ARM(SARM)患者进行基因分析,利用CONVEX和AION双管线筛选罕见致病变异,结合蛋白互作网络分析和拷贝数变异检测。结果发现:20个候选变异涉及NOTCH3、GLI2、PKD1、PKHD1等关键发育基因,其中NOTCH3变异位于表皮生长因子样结构域,可能影响配体结合;GLI2剪接位点变异破坏SHH信号通路。通过Cytohubba网络分析鉴定出PKD1、PKD2、PKHD1为核心枢纽基因,参与纤毛相关信号通路。与欧洲GWAS数据比对发现EFNA1和ISL1两个常见变异,提示跨人群遗传结构共享。本研究为SARM的分子诊断和个性化干预提供了新靶点。
今天给大家解读一篇4月发表在《iScience》上的题目为“Distinct genetic variants from the whole-exome sequencing of syndromic anorectal malformations: A cross-sectional study.”的文章。本文通过对8例综合征性肛门直肠畸形(SARMs)患者进行全外显子组测序(WES),旨在探索该疾病的遗传病因。研究使用自主研发的CONVEX和AION两个独立分析管道筛选变异,并通过IGV验证。研究共优先排序了20个候选变异,并识别出多种潜在致病基因,如NOTCH3、GLI2、PKD1等。通过与欧洲GWAS数据交叉对比,发现两个共有变异ISL1和EFNA1。网络分析揭示了PKD1、PKD2和PKHD1等是SARMs通路的枢纽基因。此外,结构变异分析在所有样本中发现了一个包含基因MIEF1的重复性CNV。研究认为这些发现为SARMs的遗传诊断和个性化干预提供了新的线索。(请持续关注我们,每天为您解读最新见刊的文献!)想薅生信资料羊毛?直接在对话框回复 “资料”,免费领取干货大礼包!包括数据集、绘图代码、图表复现、思路总结、参考文献……0代码!鼠标点点点即可轻松完成5-10分生信SCI全文复现!
不想做实验,没数据,还想要快速发表文章,没问题的!公共数据库就是我们的数据宝藏!没思路不用担心,作为专业的生信团队,我们很乐意为你们效劳,提供研究路线设计和数据挖掘分析,扫码联系我们吧!



团队成员合影(位于上海陆家嘴中心,可随时预约参观)
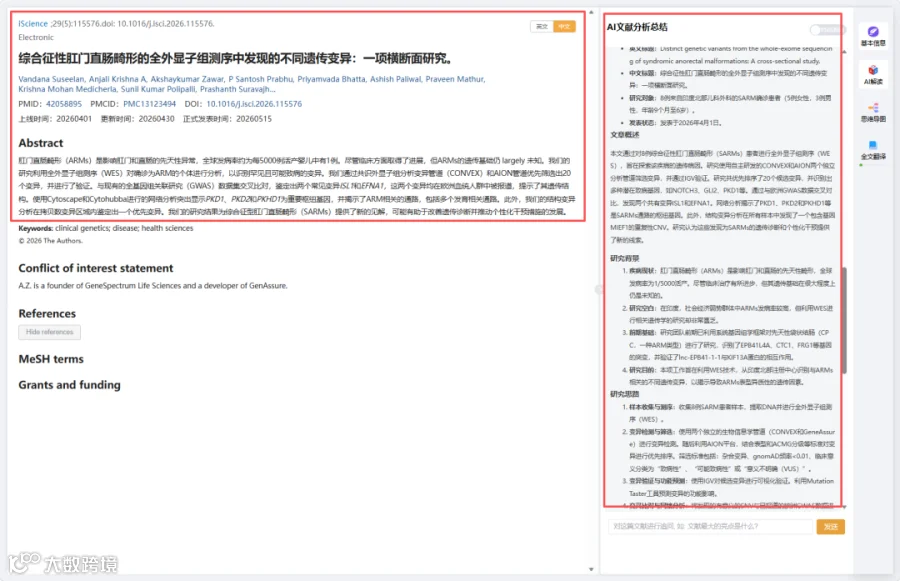
题目:《综合征性肛门直肠畸形的全外显子组测序中发现的不同遗传变异:一项横断面研究》Distinct genetic variants from the whole-exome sequencing of syndromic anorectal malformations: A cross-sectional study
发表期刊:iScience
影响因子:4.1
研究背景:
- 疾病现状
肛门直肠畸形(ARMs)是影响肛门和直肠的先天性畸形,全球发病率为1/5000活产。尽管临床治疗有所进步,但其遗传基础在很大程度上仍是未知的。
- 研究空白
在印度,社会经济弱势群体中ARMs发病率较高,但利用WES进行相关遗传学的研究却非常匮乏。
- 前期基础
研究团队前期已利用系统基因组学框架对先天性袋状结肠(CPC,一种ARM类型)进行了研究,识别了EPB41L4A、CTC1、FRG1等基因的突变,并验证了lnc-EPB41-1-1与KIF13A蛋白的相互作用。
- 研究目的
本项工作旨在利用WES技术,从印度北部注册中心识别与ARMs相关的不同遗传变异,以揭示导致ARMs表型异质性的遗传因素。
CNSknowall 平台 Pubmed+AI 快速提炼全文要点
研究思路:
- 样本收集与测序
收集8例SARM患者样本,提取DNA并进行全外显子组测序(WES)。
- 变异检测与筛选
使用两个独立的生物信息学管道(CONVEX和GeneAssure)进行变异检测。随后利用AION平台,结合表型和ACMG分级等标准对变异进行优先排序。筛选标准包括:杂合变异、gnomAD频率<0.01、临床意义分类为“致病性”、“可能致病性”或“意义不明确(VUS)”。
- 变异验证与功能预测
使用IGV对候选变异进行可视化验证。利用MutationTaster工具预测变异的功能影响。
- 交叉比对与网络分析
将发现的有意义的SNV与已报道的欧洲GWAS数据进行对比。利用Cytoscape和Cytohubba软件构建基因相互作用网络并识别枢纽基因。
- 结构变异分析
对样本进行全基因组结构变异分析,识别拷贝数变异(CNV)。
研究亮点:
-
利用WES技术识别与SARMs相关的罕见和潜在致病性变异。
-
通过多种变异检测工具,筛选并优先排序了20个不同的基因变异。
-
通过网络分析,将PKD1、PKD2和PKHD1鉴定为候选枢纽基因。
-
研究结果:
- 优先变异
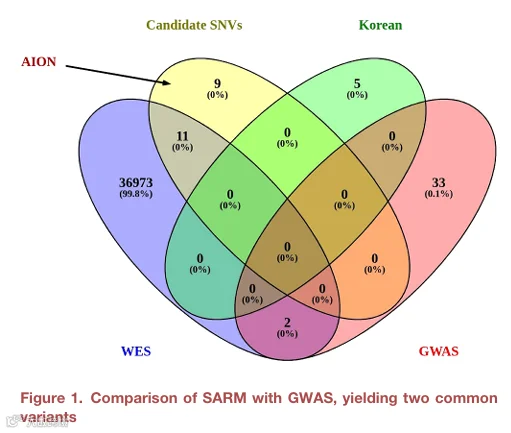
通过筛选,研究共确定了20个候选变异。其中9个由AION平台专属优先排序,其余11个由CONVEX和AION共同识别。这些变异位于与发育障碍和先天性异常相关的基因中。
- 关键基因与通路
- NOTCH3
在SARM4和SARM7样本中发现,其变异影响配体结合和受体激活的EGF样结构域,提示NOTCH通路在胚胎肠道发育中起重要作用。
- GLI2
作为SHH信号通路的关键介质,其剪接位点变异可能导致功能丧失,与后肠和泄殖腔形成相关。
- PKD1, PKD2, PKHD1
与纤毛相关信号缺陷和多囊肾病相关,被认为是SARMs的候选枢纽基因,可能通过破坏纤毛介导的信号传导影响后肠和泄殖腔发育。
- 其他基因
还包括与细胞骨架重塑(ARHGAP29)、胚胎发育(ZNF462)、钠通道功能(SCN1A)和线粒体功能(PDSS1、PYCR1)等相关的基因。
- 与欧洲GWAS的对比结果
通过交叉比对,发现两个与欧洲人群GWAS研究重叠的常见变异,分别位于EFNA1和ISL1基因。
- 结构变异分析
在所有8个样本中均发现一个位于22号染色体上的拷贝数变异(CNV),标识为nsv429523,该区域包含与线粒体动力学相关的基因MIEF1。
研究总结:
- 主要结论
本研究通过WES技术,首次在印度人群中系统性地分析了SARMs的遗传基础。识别了20个优先候选变异和多个潜在致病基因,其中PKD1、PKD2、PKHD1等纤毛相关基因和通路可能起核心作用。同时,发现了与欧洲人群共有的遗传变异(ISL1和EFNA1),提示跨人群存在共同的遗传易感性。
- 讨论与展望
- 局限性
研究样本量较小(n=8),均为独立个案(Singleton),缺乏家系共分离分析,限制了统计效力。同时,研究未深入分析非编码区和调控元件,未来有必要在更大、更多样化的队列中纳入调节和结构变异。
- 临床意义
研究为SARMs的遗传诊断提供了新靶点,有助于改善遗传咨询和开发个性化干预措施。AION平台对一些“VUS”变异的重新分类也展示了其在减少临床解读不确定性方面的潜力。
- 未来方向
需要进一步通过功能实验验证这些候选基因和变异在ARM发病中的具体作用。研究指出的POLD1等基因与结肠癌的关联,也提示了ARMs与解剖结构相关的其他疾病之间可能存在遗传通路的重叠,值得进一步探索。
结果译文:
我们使用两个独立管线——CONVEX和GeneAssure——分析了8例不同严重程度ARM患者的样本,并进一步使用AION对结果进行优先排序。在初步筛选中,我们重点关注被鉴定为杂合调用(GT=0/1)的真正候选变异,因为我们只有单例(受影响先证者,无父母测序信息)。我们根据临床意义进一步优先排序变异,特别是那些被归类为“致病性”、“可能致病性”或“意义不明确变异”的变异。此外,我们对等位基因频率设置了严格过滤,仅考虑gnomAD频率小于0.01的变异,此外还考虑了CONVEX和AION过滤的其他属性。通过这一过程,我们在所有8个样本中短列出了总共20个候选变异进行进一步分析(表1)。在CONVEX预测的20个变异中,9个由AION平台单独优先排序,其余11个在CONVEX和AION之间共享(图1)。值得注意的是,几个候选变异位于先前与发育障碍和先天性异常相关的基因中,并且所鉴定的通路被发现与ARM相关。
在SARM4(NC_000019.10;g.15179137G>C)和SARM7(NC_000019.10;g.15179052G>A和NC_000019.10;g.15185655T>C)中报道了NOTCH3的变异。由于NOTCH3在胚胎发生和组织模式形成以及肠道发育的细胞命运信号传导中的作用,它被发现与之相关。MutationTaster预测所有三个已鉴定的变异均为错义突变,定位于NOTCH3的细胞外表皮生长因子样结构域,这些结构域对配体结合和受体激活至关重要。其中两个变异NC_000019.10;g.15179137G>C和NC_000019.10;g.15179052G>A改变了保守的半胱氨酸残基,这是一种已知的因二硫键形成受损而导致EGF样结构域破坏的机制。另一个变异NC_000019.10;g.15185655T>C影响一个保守的非半胱氨酸残基,可能影响蛋白质折叠或受体-配体相互作用。这三个变异在人群数据库中罕见,功能丧失不耐受概率为0.4。NOTCH通路信号在胚胎肠道发育、上皮分化和间充质-上皮相互作用中发挥重要作用。因此,这些发现表明NOTCH3的变异可能成为ARM的候选基因。
GLI2与泄殖腔和后肠形成中的肛门直肠发育相关,因为它是SHH信号通路的关键介质。突变NC_000002.12:g.120797290G>A影响剪接位点且具有致病性。GLI2通过FGF-brachyury调节环路在中胚层模式形成中发挥重要作用。它激活brachyury和Xhox3,连接FGF和SHH通路以维持中胚层和后部特化。该变异(NC_000002.12;g.120797290G>A)位于内含子-外显子边界,可能导致基因功能改变。GLI2的剪接受体变异是一个单倍剂量不足基因,pLI高达0.97,因此该变异可能作为多基因或多因素框架内的因果或修饰等位基因。
ARHGAP29影响中线发育缺陷——口面裂,这可能是颅面和肛门直肠异常的重叠机制,其中错义突变变异(NM_004815.4: c.2224G>A)可能导致缺陷并引发ARM。我们观察到ARHGAP29变异影响Rho-GAP结构域中一个进化保守的残基。该基因表现出对功能变异的耐受不良(pLI = 0.99),对错义变化具有潜在敏感性。在颅面和胚胎发育的组织重塑过程中,ARHGAP29调节RhoA介导的细胞骨架动力学和上皮形态发生。ARHGAP29功能障碍可能作为共享的中线发育通路的一部分,促成ARM易感性。类似地,ZNF462的变异是一种锌指转录因子,在胚胎发育和染色质重塑中发挥作用,被发现与Weiss-Kruszka综合征相关,该综合征包括心脏、骨骼和泌尿生殖系统异常。我们的研究鉴定出ZNF462的一个错义变异NC_000009.12:g.106972095C>G(p.Pro2173Arg),可能导致发育综合征。该变异在人群数据库中极为罕见,pLI=1,为单倍剂量不足,该基因相关的变异被预测为有害,且其存在于一个对胚胎模式形成至关重要的基因中,提示它可能与其他遗传变化组合增加ARM风险。
SCN1A是一个钠通道基因,与癫痫和神经发育相关,具有综合征背景。在我们的研究中,NC_000002.12:g.166046808T>C错义变异因其功能相关性而被优先考虑。虽然该变异在ClinVar中被归类为“VUS”,但其极低的人群频率、有害预测和基因水平的不耐受性表明其对ARM可能存在功能影响。
PYCR1和PDSS1是代谢基因,我们观察到其变异与ARM发病相关。PDSS1中的无义变异NC_000010.11:g.26723857C>T引入了一个提前终止密码子,预计会触发无义介导的mRNA衰变,导致功能性蛋白缺失。该变异在ClinVar中被归类为“致病性”,并与线粒体功能障碍相关的多系统疾病相关联。PDSS1编码辅酶Q10生物合成途径中的一种必需酶,功能丧失变异影响线粒体氧化磷酸化,导致发育异常。早期胚胎发生过程中的线粒体功能障碍在复杂的综合征病例中很重要,ARM可能作为次要特征出现。PYCR1导致皮肤松弛综合征,包括从发育障碍到结缔组织异常的多种缺陷。在PYCR1基因中鉴定出一个错义变异NC_000017.11:g.81935368G>A,该变异在所有评估的转录本中均被预测为有害,高置信度树投票为95%-98%。该变异在人群数据库中罕见,氨基酸替换支持潜在的功能影响。PYCR1在脯氨酸生物合成中发挥重要作用,该通路对胶原完整性、细胞氧化还原平衡和正常胚胎发育至关重要。这些过程的破坏可能产生复杂的综合征表型,其中ARM可作为次要表现出现。
PKHD1和PKD1已知与纤毛相关信号缺陷有关,参与先天性异常如多囊肾病,这是ARM相关异常。在PKHD1中鉴定出的错义变异(NC_000006.12:g.51746863T>C)被MutationTaster预测为跨多个转录本有害。这影响fibrocystin蛋白中一个保守的氨基酸残基。PKHD1编码一种大的纤毛蛋白,参与上皮形态发生和平面细胞极性信号传导,这对胚胎发生过程中的组织重塑至关重要。由于纤毛介导的信号在胃肠和泄殖腔发育中很重要,该变异可能易致ARM。我们在SARM4中鉴定了两个PKD1的错义变异。变异NC_000016.10:g.2103386C>T被预测为跨多个转录本有害,它导致氨基酸替换而不影响蛋白质长度或剪接位点。该变异在人群数据库中频率较低,功能丧失不耐受。纤毛依赖性通路破坏可导致后肠和泄殖腔发育异常。这表明该变异可能作为导致ARM的遗传修饰因子。另一个PKD1错义变异NC_000016.10:g.2103820C>T也被预测为有害,人群频率低,pLI=1。
CCDC50基因突变导致听力损失。由于这是ARM的相关异常,我们考虑了优先排序导致蛋白提前终止(p.Ser276Ter)的变异NC_000003.12:g.191375440C>G。终止增益变异NC_000003.12:g.191375440C>G基于这种重叠被优先排序。虽然这些基因先前未在ARM中报道,但GeneCards GTEx显示它们在结肠和胃肠组织中表达,提示可能影响SARM相关发育通路。在变异水平上,NC_000003.12:g.191375440C>G变化引入了一个提前终止密码子,预计会发生NMD。OSMR,一种制瘤素M受体,在影响上皮和间充质发育的细胞因子信号传导中发挥作用,其携带一个终止增益突变NC_000005.10:g.38904479C>T,导致蛋白功能丧失并终止(p.Gln421Ter)。该无义变异被归类为有害,预计会发生NMD并导致蛋白质截短,丢失所有胞外纤连蛋白III型结构域、跨膜区和胞内信号基序。OSMR是IL-6家族细胞因子信号传导中的关键受体,介导JAK-STAT依赖性通路,这对上皮-间充质串扰和胚胎组织模式形成至关重要。这些信号的破坏可能影响后肠和泄殖腔发育,可能解释OSMR功能丧失在ARM中的作用。
GUCY2C,一种鸟苷酸环化酶,调节肠道上皮并可能导致肠道畸形。在该基因中鉴定出NC_000012.12:g.14621060G>A,导致错义突变p.Arg920Cys。虽然该变异在脊椎动物物种中高度保守,但它位于鸟苷酸环化酶催化结构域内,提示其可能降低酶活性。由于cGMP介导的信号在肠道上皮成熟和肠道形态发生中发挥重要作用,GUCY2C的破坏可能导致ARM发病。TYR主要参与黑色素生物合成,与色素沉着障碍相关,但它在神经嵴衍生物中表达,提示错义变异NC_000011.10:g.89284805C>T具有潜在的综合征重叠。作为一个潜在有害的变异,TYR先前未报道与ARM相关,但神经嵴来源的细胞有助于黑色素细胞发育和后肠形态发生。因此,这些发育通路的破坏可能导致SARM,因为ARM可能源于胚胎发育破坏。
PSAP(prosaposin)对溶酶体功能和神经发育很重要,与SARM相关。在PSAP基因(chr10:71819830T>G)中发现了一个罕见的错义变异,该变异与鞘脂代谢缺陷引起的疾病相关。受影响的氨基酸位于saposin B结构域内,该结构域对正常的蛋白功能很重要,提示这一变化可能破坏蛋白质结构。此外,PSAP是一个高度保守且功能重要的基因,pLI=0.99。溶酶体鞘脂代谢对正常胚胎发育和组织形成至关重要;因此,该变异可能增加ARM的易感性。
ZEB1是一个转录抑制因子,促进上皮-间充质转化,在发育综合征中发挥作用并可能影响后肠形态发生。在我们的样本中,ZEB1变异NC_000010.11:g.31514613C>T导致错义突变,可能是ARM的致病原因。虽然它被预测为在所有受影响的转录本中有害,并且在人群数据库中极为罕见,pLI=0.9,但该基因对功能破坏不耐受。MIEF1是一个线粒体动力学调节因子,通过与DNM1L相互作用调节线粒体分裂。我们在研究中鉴定出MIEF1的一个罕见错义变异NC_000022.11:g.39513632G>A,它在人群数据库中频率极低,被MutationTaster预测为跨多个转录本有害。改变的残基位于与DNM1L相互作用的一个保守区域内,DNM1L是调节线粒体分裂的关键蛋白。早期胚胎发生过程中线粒体动力学受损可能导致组织发育和细胞分化异常。
我们短列出的变异(表1和表S1)揭示了一个关于C和G碱基突变位点的有趣观察。先前的研究指出,罕见孟德尔疾病中的生殖系变异表现出与群体变异相似的一般偏倚,其中转换显著多于颠换——在生殖系中大约频繁3-4倍。具体来说,甲基化CpG位点的C→T(以及互补的G→A)变化在突变谱中占主导地位。例如,ExAC/gnomAD等大型数据集表明,所有可能的CpG→TpG转换中约有60%-80%发生在人群中,而可能的颠换仅占一小部分。类似地,特定隐性遗传病基因中近一半的错义突变发生在CpG位点,其中约74%是C→T/G→A转换。这些CpG→TpG转换源于自发的5mC脱氨作用,对应于众所周知的生殖系特征SBS1。相比之下,其他环境如A/T位点或非CpG变化显示出低得多的突变性。总之,罕见的常染色体隐性遗传病等位基因富含CpG环境中的G/C→A/T转换。这种富集反映了标准的生殖系突变偏倚。
我们尝试比较CONVEX和AION的分类,特别是ClinVar未分类的变异。例如,AION通过整合多个注释和基于群体的矩阵,成功鉴定出GLI2和OSMR中的致病变异,将其归类为“致病性”。此外,ARHGAP29、PKHD1、NOTCH3、ZNF462和MIEF1中的变异被指定为“可能致病性”。通过对这些变异进行分类,AION预测了可能在ARM中起关键作用的候选基因,超越了ClinVar当前的局限性。例如,SARM7中的一个NOTCH3基因变异(NC_000019.10:g.15179052G>A),ClinVar将其归类为致病性解释冲突,但AION将其标记为“致病性”。鉴于NOTCH3在发育信号传导中的作用,这提示它可能参与SARM,而AION可能有助于解决这种解释上的不确定性。此外,ClinVar归类为“VUS”的几个变异(NC_000002.12:g.16604680T>C、NC_000017.11:g.81935368G>A、NC_000003.12:g.191375440C>G和NC_000012.12:g.14621060G>A)被AION标记为“致病性”或“可能致病性”。这突显了AION在减少不确定性和为这些变异提供临床相关性方面的准确性。类似地,AION将一些ClinVar VUS重新分类为“良性”或“可能良性”,展示了其降级相关变异和澄清不确定分类的能力。然而,也存在一些例外。例如,变异NC_000007.14:g.6006003T>C在ClinVar中被认为“可能良性”,但被AION预测为“致病性”。相反,ClinVar中归类为“可能致病性”或“致病性”的变异NC_000011.10:g.89284805C>T被AION确认为“可能致病性”,表明良好的一致性。另一方面,变异NC_000010.11:g.71819830T>G在ClinVar中被标记为“致病性”或“可能致病性”,但被AION标记为“VUS”,表明注释存在细微差异。此外,PDK1变异最初在ClinVar中被归类为“VUS”,被AION重新分类为“良性”。相反,AION将CCDC50和TYR变异分类为“致病性”。在9个变异中,有一个(NC_000007.14:g.6006003T>C)在ClinVar中被预测为“可能良性”,但被AION预测为“致病性”,AION变异评分为0.94,可能提示致病性。我们还将我们的候选变异与先前在韩国年龄相关性黄斑变性队列研究中报道的变异进行了比较,但未发现共同变异。这种差异可能源于评估变异的设计和方法学差异,此外仍有重要变异有待诊断评估。
我们还识别出可能导致ARM的潜在遗传因素,并确定了两个值得进一步研究的基因:CABP4和POLD1。CABP4基因编码一种对肾突触信号传导至关重要的钙结合蛋白,主要与视网膜营养不良相关。然而,它在胎儿发育过程中神经信号传导中的作用提示,突变可能影响对肛门直肠发育至关重要的神经嵴细胞。虽然确切机制尚不清楚,但CABP4突变导致的神经分化过程破坏可能引起ARM。另一方面,POLD1基因编码DNA聚合酶δ的一个亚基,对DNA复制和修复至关重要。POLD1突变与多种癌症以及聚合酶校正相关息肉病综合征(增加结直肠癌风险)等相关。POLD1突变可能影响下消化道发育,导致ARM。由POLD1突变引起的DNA修复机制损伤可能导致胎儿发育过程中组织分化异常,导致ARM或相关疾病。
3.SARM与欧洲GWAS的比较产生了两个常见变异
在将我们的SARM队列与先前报道的GWAS进行交叉比对时,我们鉴定了两个重叠的感兴趣变异(表2和图1)。第一个是EFNA1中的杂合错义变异(NC_000001.11:g.155133751A>T),在我们的7个样本中观察到。该变异最初由Mingardo等人在其GWAS中报道,显示与膀胱外翻有强烈的统计学关联,p值为4×10⁻¹⁰。第二个重叠变异位于ISL1(NC_000005.10:g.51389671A>G),在我们的5个样本中鉴定出,先前由Draaken等人在2015年报道。这些变异同时出现在我们的数据和早期的欧洲GWAS中,表明ARM在不同人群中可能存在共享的遗传背景。一项对450名ARM个体的全基因组拷贝数变异分析鉴定出4个显微镜下可见和9个亚显微CNV(del2p13.2、del4p16.2、del7q31.33、del9p24.1、del16q12.1、del18q32、del22q11.21、dup2p13.2和dup17q12)。研究者经过一些过滤后优先考虑了这些染色体异常。基于胚胎表达和功能,FOXK2、LPP和SALL3被提出作为该研究中ARM的候选基因。
在我们的流程中,这些变异未达到过滤标准,该标准要求gnomAD等位基因频率低于0.01,且仅包含“致病性”、“可能致病性”和“VUS”变异。此外,我们将分析限制在SARM,而GWAS也关注其他类型的ARM。尽管如此,我们在放宽过滤条件时发现了一些共同变异,这促使我们通过Integrative Genomics Viewer进一步验证(图S1-S18)。
另外,我们观察到NC_000013.11:g.20189511C>T与SARM1相关,这是由于AION的HPO映射所致。GJB2主要已知引起听力损失;然而,变化NC_000013.11:g.20189511C>T引入了一个终止增益突变(p.Trp24Ter)。这一点很重要,因为ARM可与其他异常如猫眼综合征(包括眼缺损、耳异常和ARM)同时发生。先前一项研究报道,发育不良的耳朵、听力损失和ARM共同发生,支持听力相关基因的致病变异提示与ARM存在可能的重叠遗传关联的证据。虽然GJB2与ARM无关,但我们认为这是AION不准确映射HPO术语的一个局限性。MutationTaster分析支持GJB2变异的致病性。通过该突变在氨基酸位置24引入了一个提前终止密码子,导致蛋白质截短。这种截短可能破坏GJB2的功能,因为GJB2编码连接蛋白26,一个间隙连接蛋白。连接蛋白形成的间隙连接通道在细胞间通讯中至关重要,对正常组织发育和稳态不可或缺。间隙连接功能障碍可导致多系统发育缺陷,包括听力损失和潜在的肛门直肠异常。这提示GJB2可能通过多效性或通过破坏共享的发育通路间接导致ARM发病。
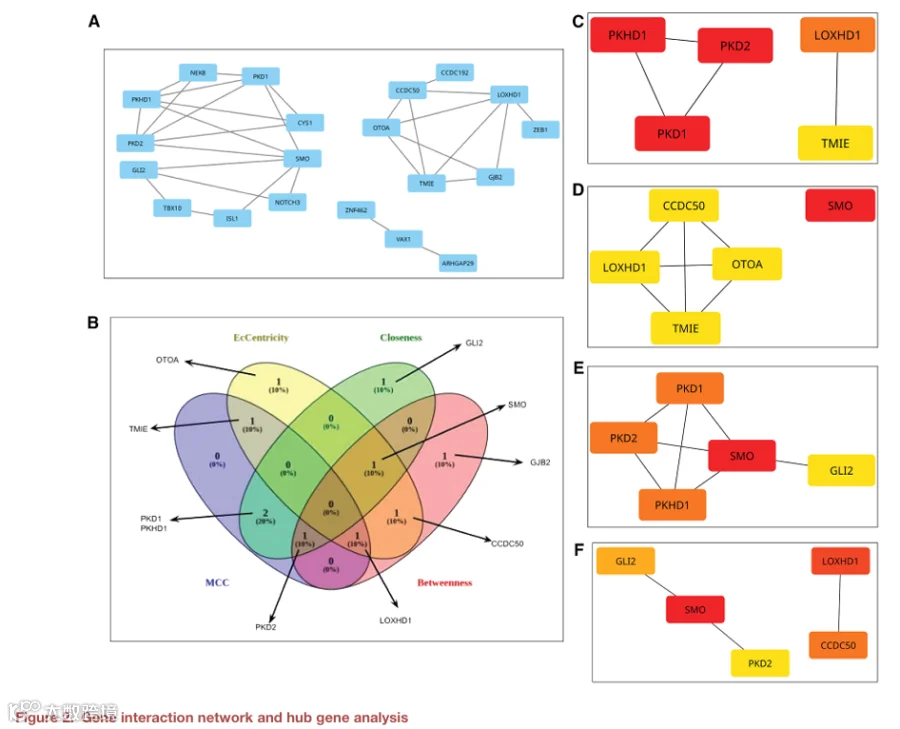
为了更全面地了解ARM的遗传结构,我们进行了网络分析以确定中心基因(图2A)。使用STRING数据库构建基因-基因相互作用网络,然后导入Cytoscape并使用Cytohubba插件通过四种中心性度量(MCC、偏心度、紧密度和介数)进行排名。排名靠前的基因总结于表3。值得注意的是,根据这些度量,PKD1、PKD2和PKHD1始终被确定为重要的中心基因,其中PKD1被鉴定为网络中最中心的节点(图2C-F,表3)。这一发现在ARM背景下具有特别的意义,因为这些基因是多囊肾病的关键基因,而多囊肾病是一种以肾脏囊肿为特征的疾病。已知肾脏异常是VACTERL联合征中ARM最常见的相关异常之一,这是一个重要的临床关联。此外,纤毛和纤毛信号在肾脏和肠道发育中都起着至关重要的作用。SHH和WNT信号通路在脊柱、肾脏和泄殖腔发育中不可或缺,依赖于功能性纤毛。这些通路中的缺陷可能同时导致肾脏和肛门直肠异常,正如GLI2(SHH通路的一个关键转录因子)的变异被确定为ARM易感性相关一样。此外,PKHD1编码fibrocystin,一种定位于肾上皮细胞初级纤毛的蛋白质,突显了其更广泛的发育作用,超越了肾脏形态发生。我们在PKD1基因中鉴定了两个错义变异(NC_000016.10:g.2103386C>T和NC_000016.10:g.2103820C>T),目前在ClinVar中归类为“VUS”。因此,这些变异对肛门直肠发育的临床影响仍不清楚,因为当前文献中的证据不足或冲突。由于PKD1对纤毛相关信号传导至关重要,这些变异代表了进一步研究ARM遗传结构的候选对象。
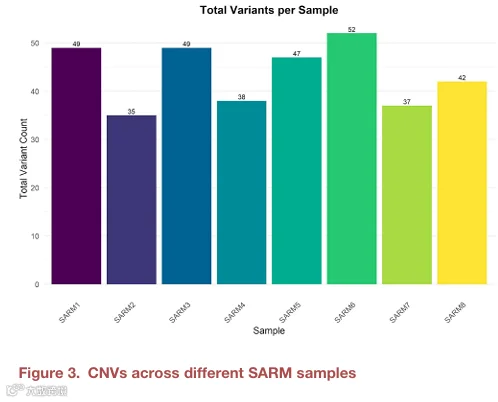
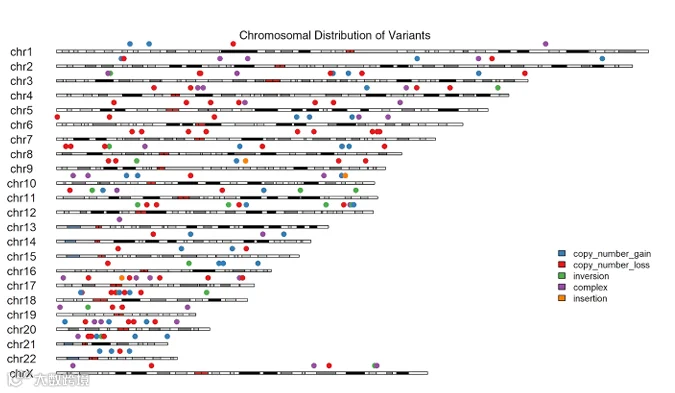
我们分析了所有8个样本的结构变异,共鉴定出170个变异(图3和表S2)。其中几个变异在多个样本中共享。值得注意的是,一个CNV——nsv429523——在所有8个样本中均观察到(图4)。该CNV位于22号染色体上(23155487-50796027),该区域包含我们优先考虑的基因之一MIEF1。MIEF1编码一种与线粒体动力学相关的蛋白质,已知线粒体功能的改变会导致发育异常。该CNV在所有样本中的一致存在提示线粒体影响可能在ARM的发育中起作用。
我们还试图检查Genome India Project中变异的排列组合。总的来说,从GIP中已发布的替代等位基因频率列表来看,我们发现存在于ARM但不存在于gnomAD和GIP等大型健康人群数据库中的变异,表明它们确实是罕见的,因此是致病性的强候选者。值得注意的是,我们选择的ARM候选在很大程度上与VACTERL联合征一致,其中六个在GIP中不存在的变异可能被优先作为致病变异,因为它们可能具有高pLI,提示该区域的功能变异是ARM表现的基础。
更多结果和补充图表:doi: 10.1016/j.isci.2026.115576
长按二维码关注我们,用最短的时间和最高的效率学习更多生信思路!
扫描上方二维码或登录平台官网后添加CNSknowall客服微信咨询!官网地址:https://cnsknowall.com
CNSknowall:24年最新问世的遥遥领先的颠覆性科研数据(0代码生信+统计学)分析平台,同时含有机制图模块(原创3000多素材和机制图模板)+AI一键生成高质量比国自然标书初稿+汉化版Pubmed融合Deepseek高效筛选目标文献同时一键提炼全文核心创新点+SCI文献例句/语料检索模块+全文翻译+文献求助+图片查重+期刊查询+OPenAI官方GPT接口,>500款CNS级别图表皆可一秒内一键出图,登录即秒变数据分析大神,体验前所未有的便捷数据分析之旅,开启科研天骄之路!
可向下滑动发掘更多科研秘籍!