
病毒防治的世界难题被攻克!内生菌防治病毒被证实!
-----番茄斑萎病毒防治达到95.73%!
国际顶尖学术期刊《害虫管理科学》(英国,1 区 TOP 期刊1)近日发表了云南农业大学Rizwan Khan、何月秋教授等的学术论文《内生芽孢杆菌在番茄斑萎病毒可持续治理及产量提升的应用》,研究报告了五株芽孢杆菌对番茄斑萎病毒的防治效果分别达到 91.21%、64.92%、 76.43%、94.07% 和 95.73%,而且,内生菌还显著改善了生长参数, 增加了茎和根的长度、叶片数量、叶面积和生物量。此外,内生菌的应用激活了关键的抗氧化酶(多酚氧化酶、过氧化物酶、超氧化物歧化酶和几丁质酶), 同时降低了应激和防御相关指标(丙二醛、苯丙氨酸解氨酶、总酚和β-1,3-葡聚糖酶),从而保护植物免受病毒引起的损害,为病毒病的防治提供了新的途径。
这是有文献记录以来首次发表内生菌防治番茄斑萎病毒病的研究论文!攻克了只能“抗”病毒病,而不能“治”病毒病的世界难题!

内生芽孢杆菌在番茄斑萎病毒可持续治理及
Rizwan Khan, Pengbo He, Xiaojiao Chen, Pengfei He, Ayesha Ahmed, Yixin Wu, Guowen Tang, Ping Tang, Xingyu Li, Shahzad Munir* and Yueqiu He*
摘要
背景:番茄斑萎病毒(TSWV)属于托孢病毒属, 可侵染 1000 多种植物, 其中包括重要农作物, 而传统的防治方法往往效果不 佳。本研究旨在探究枯草芽孢杆菌和解淀粉芽孢杆菌在降低番茄斑萎病毒的感染率、促进植物生长以及增强植物抗性方面对本 氏烟的作用效果。其目的是评估芽孢杆菌作为可持续生物防治手段的潜力, 为农业中番茄斑萎病毒病害的管理提供一种环保的 解决方案。
结果:在此,我们报告了 15 株芽孢杆菌中 5 株(枯草芽孢杆菌 L1-21 和解淀粉芽孢杆菌 DJB5、YN48、YN28、Mg6)在半叶试 验中显著降低了本氏烟叶片中番茄斑萎病毒(TSWV)的拷贝数。在温室试验中, 菌株 DJB5、YN48 和 Mg6 分别使每克叶片中 的 TSWV 拷贝数降低了 75.7%、83.6% 和 88.2%,生物防治效果分别为 91.2%、94. 1% 和 95.7%。所有菌株均能持续缓解 TSWV 症状, 减轻病情严重程度, 并降低接种后 21 天的病情进展曲线下面积(AUDPC)。此外, 这些菌株还能促进植物生长参数,
包括茎长、根长、叶片数量、面积和生物量。在受感染植物中应用内生菌可激活抗氧化防御酶,提高多酚氧化酶(PPO)、过 氧化物酶(POD)、超氧化物歧化酶( SOD)和几丁质酶的活性。然而, 随着叶片中TSWV 感染的减轻, 防御相关酶如丙二醛 (MDA)、过氧化氢酶(CAT)、苯丙氨酸解氨酶(PAL)、总酚和β-1,3-葡聚糖酶的活性降低。
结论:我们的研究结果表明,枯草芽孢杆菌(B. amyloliquefaciens)分离株 DJB5、YN48 和 Mg6 能够通过激活植物防御机制、 降低病毒载量、减轻番茄斑萎病毒(TSWV)症状以及促进植物生长来有效控制番茄斑萎病毒。
⑥2024 化学工业学会。
有关支持信息可在本文的网络版中找到。
关键词:生物防治;内生菌;烟草;病毒;防御激活
1 简介
植物病毒是导致巨大经济损失的新兴病原体之一,据估计每年 造成的损失达 300 亿美元。番茄斑萎病毒(TSWV)属于布尼 亚病毒科的托斯波病毒属,是造成农业损失最严重的十大植物 病毒之一, 每年造成的损失估计超过 10 亿美元。TSWV 由蓟 马以持久增殖的方式传播, 也可通过机械接种传播。TSWV 具有三段式基因组, 由小 RNA 段( S)、 中 RNA 段(M)和 大 RNA 段(L)组成。S 段编码一种非结构(NS)蛋白, 该 蛋白具有 RNA 干扰抑制作用, 可抵御植物的先天免疫系统。 防御系统的激活会通过抗性基因(Tsw)介导细胞死亡。因此, TSWV 能够轻易克服所有作物的抗性,使植物易受感染。
番茄斑萎病毒(TSWV)的全球流行率正在上升, 已有超 过 80 个科的 1000 多种植物被报道感染该病毒,许多农作物也因此受到影响。11, 12 开发抗病毒作物品种被认为是管理病毒的更好策略, 同时结合综合病害管理 方法。13 杀虫剂曾被用于成功控制持久性和半持久性植物病毒。 然而,这种方法会带来环境污染、害虫抗药性以及由于媒介种 群遗传变化导致的病原体重新出现等问题。14 但是,控制昆虫 媒介, 尤其是蓟马和螨虫, 15 并非总是可行的, 而且往往会对 环境造成重大影响和经济影响。16 系统获得性抗性( SAR)激活剂在病毒控制中 至关重要, 对于理解植物抗病性也必不可少。 SAR 是植物 的一种广谱免疫反应 , 通常由水杨酸( SA) 及其类似物 (如乙磷铝)触发。17 这些激活剂通过使植物为迅速的防御 反应做好准备, 并增强病程相关蛋白的表达, 从而增强植 物对病毒的抵抗力, 保护植物免受各种病毒病原体的侵害。 这增强了植物对病原体感染的抵抗力。
内生菌最近已成为增强系统获得抗性( SAR)激活的有前 景的工具。18 内生菌能激活植物的防御途径, 并有助于更可 持续和环保地管理植物病原体疾病。 因此, 研究内生菌作 为 SAR 激活剂, 为管理病毒性疾病提供了一种综合方法, 同时减少对化学治疗的依赖。 内生菌还能激活植物的防御 系统, 并与植物中存在的其他有益微生物合作, 抵御入侵 的病原体。19,20 此外, 内生菌还能诱导植物对植物病原体微 生物产生 SAR;大多数相关研究针对的是真菌21 和细菌, 22 而针对烟草花叶病毒(TMV)和黄瓜花叶病毒(CMV)等 病毒的研究则较少。23内生菌会分泌多种具有生物活性的次生化合物, 包括萜类 化合物、 酚类化合物、 肽和肽聚糖、 生物碱、铁载体、 细 菌素和脂肽 。24,25 一些有益细菌 , 如植物根际促生菌(PGPR) , 也能定殖于植物的各个部位, 通过提供营养物 质和维生素 B 来促进植物生长。26 植物防御相关基因(如 NPR1、PR、WRKY、LOX 和 PDF) 的表达和调控与各种 激素和抗氧化剂的产生密切相关。27 然而, 内生菌与病毒的 相互作用比与其他植物病原体的相互作用更为复杂, 因为 它除了激活水杨酸信号通路外, 还会激活茉莉酸信号通路。28,29
由于内生菌能直接或间接激活植物的防御途径,我们认为 减轻植物中的病毒感染可能为农业中植物病毒的控制提供 一种环保的综合病害管理(IDM) 方法。 本研究旨在探究 芽孢杆菌内生菌在可持续控制番茄斑萎病毒(TSWV) 方 面的潜力及其对生长参数的影响。 我们还评估了内生菌在 应对TSWV 感染时激活防御相关酶的效力, 突出了它们在 增强植物抗逆性和生产力方面的作用。 在此, 我们报告了 通过芽孢杆菌内生菌实现病毒生物防治的一项重大进展, 证明了它们作为对抗 TSWV 的强效抗病毒剂的有效性。在 评估的五种内生菌中, 有三种表现出显著的抑制病毒效果, 表明它们具有作为生物防治抗病毒剂的潜力。 这些结果标 志着植物病毒管理方面的重要进展, 因为我们证明了特定 内生菌的抗病毒活性, 并有助于为传统的化学处理方法创 造更环保的替代品。 这一发现为在综合病虫害防治方法中 研究内生菌开辟了新的途径。
2 材料与方法
2.1 植物材料和病毒接种体
本氏烟种子在湿度为 70% 的泥炭土中发芽, 并在温室条件 下黑暗培养 3 天。 幼苗移栽至温室的大花盆中, 培养温度 为 24 至 28 摄氏度(昼/夜) , 光照周期为 16 小时光照/8 小 时黑暗。 用于农杆菌浸染实验的是 4 至 6 周龄、 叶片完全展开 的本氏烟植株 。 本研究采用文献中所述的来自莴笋(Lactuca sativa) 的 番 茄 斑 萎 病 毒 (TSWV) 分 离 株(TSWV-LE)11 来实现植物的自然感染。按照 Aguilar 等人 30 所描述的技术, 通过汁液法接种 TSWV, 并在本氏烟上维 持该病毒。从感染植株上采集叶片, 并在 -80摄氏度下长期 保存, 以供本实验进一步研究使用。
2.2 TSWV 对烟草弯胞夜蛾的农杆菌介导侵染
番茄斑萎病毒(TSWV) 的 S、M 和 L 段以及 P19-HcPro-b (P19) 和 GFP-TSWV 构建体均通过农杆菌菌株 GV3101克隆, 由云南农业大学病毒学实验室(中国昆明) 提供。
将每个构建体在添加了抗生素(50 μg mL-1 卡那霉素和 20 μ g mL-1 利福平) 的 Luria Bertani(LB)培养基中培养, 以 获得新鲜的克隆培养物。按照农杆菌侵染法, 用 1 mL 无针 头注射器将 4 至 6 周龄的本氏烟(N. benthamiana)完全展 开的叶片进行侵染, 然后在室温下培养直至出现症状。
在植物出现番茄斑萎病毒(TSWV)症状后, 使用广州 麦格生物科技有限公司生产的 HiPure HP 植物 RNA 小型试 剂盒, 按照制造商的说明从冷冻叶片中提取总 RNA。简而 言之 , 使用中国百迈生物技术有限公司的 iScience 试剂盒 (All-in-One First Strand Synthesis Master-mix) 将 RNA 反 转录为互补 DNA(cDNA)。 在 1%琼脂糖凝胶上检查
RNA 和 cDNA 的质量, 并使用美国赛默飞世尔科技公司的 Thermo Scientific Nano-Drop 2000 分光光度计评估其纯度。
RNA/cDNA 样品在 -80。C 下保存以备后续研究。 使用美国 应用生物系统公司的聚合酶链式反应(PCR)试剂盒, 在 20 μL 的反应体系中 , 通过使用 TSWV 特异性引物对
(TSW. 1;5'-0-TCTGGTAGCATTCAACTTCAA-3'0 和TSW.25'-0-GTTTCACTGTAATGTTCCATAG-3'0) 扩 增 628 bp 的产物来验证感染叶片样本中的构建体( S = N 蛋 白)。反应体系包含 10 μL 的主混合液、 1 μL 的每种引物 ( 100 nM))、7 μL 的双蒸水 (ddH2O) 和 1 μL 的 cDNA 模板。 优化后的 PCR 条件包括 :94。C 初始变性 4 分钟, 随后进行 40 个循环 , 每个循环 94。C 45 秒 , 45。C 退火 45 秒 , 72。C 延伸 1 分钟, 最后在 72。C 延伸 10 分钟。 最终的 PCR 产物 在 1% 琼脂糖凝胶上通过凝胶电泳进行验证, 并使用 Tanon Mini-Space 1000 凝胶成像系统进行观察。 随后 , 扩增的PCR 产物使用 Sanger 测序仪(上海勤科生物科技有限公司, 中国上海)进行测序。 获得的 TSWV-N 序列与从美国国家 生物技术信息中心(NCBI)GenBank 中检索到的序列使用 BLAST 进 行 比 较 , 并 使 用 MEGA-X 软 件 通 过 邻 接 法 (Neighbor-Joining)构建系统发育树, 进行 1000 次自举检 验。
2.3 质粒构建及 qPCR 标准曲线绘制
使用 Sangon Biotech 公司(中国上海) 的 SanPrep Column DNA 凝胶提取试剂盒, 按照制造商的说明从琼脂糖凝胶中 提取扩增的 PCR 产物。然后, 利用克隆试剂盒(货号 6013; Takara 公司, 日本滋贺县) 将纯化的 PCR 产物(628 个碱 基对)连接到pMD19-T 载体(2692 个碱基对)上。 随后, 在 LB 培养基上筛选阳性转化子(白色菌落)。
培养基中添加了 X-Gal(5-溴-4-氯-3-吲哚基-β-D-半乳糖苷),
以便进行蓝白筛选。从 LB 培养基中挑取白色菌落, 并使 用 PCR 热循环仪(Applied Biosystems 公司) 进行菌落 PCR, 以确认目标插入片段已成功转入载体。在 2%琼脂糖 凝胶上确认后,对 TSWV-N 插入片段进一步进行测序验证, 并使用BLASTn 与从 NCBI 数据库中检索到的序列进行同 源性比对。
采用 Applied Biosystems32 的方法生成了定量 PCR(qPCR)标 准曲线, 使用特异性引物对(TSWV-F2;5'-0-GCTTGTTG AGGAAACTGGGAATT-3'0 和 TSWV-R2;5'-
AGCCTCACAGACTTTGCATCATC-3)对样本中的 TSWV- N 绝对拷贝数进行定量。通过克隆 117 个碱基对的 TSWV- N 蛋白产物生成标准曲线。31 因此, 使用含有 30 至 300 万 份每微升 TSWV-N 病原体拷贝数的六种连续稀释液制备 TSWV-N 质粒。将 PCR 反应(20 微升)设置为每份 qPCR 反应中加入 5 微升质粒 DNA。
2.4 生物防治试验
2.4.1 半叶测定法和生物防治剂
总共从中国昆明云南农业大学分子植物病理学实验室获取了 15 株内生菌。将这些菌株接种于 LB 培养基上, 制备新 鲜培养物, 然后在 37。C 下培养 48 小时。从每个菌株中选 取单菌落, 通过反复划线进行纯化, 并在 -80。C 下用 50% 甘油保存。为了筛选对番茄斑萎病毒(TSWV)具有生物 防治作用的菌株, 将每种细菌接种到 LB 液体培养基中,在 37。C 下培养 48 小时。培养结束后, 将细菌以 1660×g 的转速离心 10 分钟。弃去管中的上清液, 将沉淀悬浮于灭 菌的 ddH2O 中, 并涡旋至均匀。在半叶测定法中应用内生 菌之前 , 将细菌悬液调整至 10-8 CFU/mL(OD600 = 1)。 然后向细菌悬液中加入吐温 -20(0.1 μL/mL), 并充分混 合。
最初,按照第 2.1 节所述, 采用文献中描述的汁液接 种法, 用 TSWV-LE 分离株对 4 至 6 周龄且叶片完全展开 的 N. benthamiana 植株进行接种。30 在半叶试验前, 通过 qPCR 从受感染植株的叶片中确认了 TWSV-N 的存在。33 在半叶试验中,将一片完整的叶子切成两半;其中一半用每毫升含 1 × 10-8 CFU 的细菌内生菌悬液处理, 并置于培 养皿( 180 毫米) 中, 另一半用灭菌的 ddH2O 处理作为阴 性对照。实验中每种处理均重复五次, 每次重复包含五片 叶子。然后将培养皿在 28。C 下培养 7 天, 随后按照第 2.2 节所述提取 RNA 并合成 cDNA。通过 qPCR 定量检测病原 体拷贝数的减少, 并根据生成的 TSWV-N 标准曲线进行计算。 在实时荧光定量 PCR 系统(Applied Biosystems 公司 的 Step One Real-Time PCR 系统) 中, 反应总体积为 20 μ L, 其中包括 10 μL 的 PCR 缓冲液( SYBR® Premix Ex Tag, Takara 公司) , 每对引物(TSWV-F1/TSWV-F2)各 0.4 μ L, 0.4 μL 的 ROX, 6.8 μL 经二乙基焦碳酸酯(DEPC)处 理的灭菌水 ddH2O, 以及 2 μL 的反转录所得的 cDNA。反 应条件为:95 。C 预变性 1 分钟, 然后进行 45 个循环, 每 个循环包括 95 。C 变性 15 秒、59 。C 退火 15 秒和 72 。C 延 伸 45 秒。根据 qPCR 中测得的循环阈值(CT)值, 并参 照对照和内生菌处理叶片的标准曲线, 计算出每克叶片中 病原体的拷贝数。
2.4.2 内生芽孢杆菌对番茄斑萎病毒的防治效果
这些植物的培育方式如第 2.1 节所述, 在温室中进行生物 防治实验。从半叶测定中筛选出的两种内生菌(枯草芽孢 杆菌 DJB5、YN28、YN48 和 Mg6 以及解淀粉芽孢杆菌 L1- 21)的五个菌株, 根据对病原体的显著抑制效果, 接种在 LB 培养基上, 具体方法如前所述。首先, 除了阴性健康对 照(H-CK)外, 用汁液接种法将番茄斑萎病毒(TSWV) 接种到 4 至 6 周龄、 叶片完全展开的烟草弯胞菌(N.
benthamiana)植株上。在接种植株出现 TSWV 症状后, 除 了阳性感染对照(I-CK)和 H-CK 外, 每株植物喷施 15 毫 升细菌悬液( 108 CFU 毫升-1)。
该实验采用随机区组设计(RCBD), 重复两次, 每 个处理重复四次, 每次重复包含三株植物。实验期间每两 天灌溉一次, 实验过程中未添加肥料。在接种后 0、7、 14 和 21 天(DPI)分别记录病原体每克减少量、病情严重程 度以及病情进展曲线下面积(AUDPC)。此外, 还记录了 植物生长促进参数, 包括株高(厘米)、每株叶片数、叶 面积(平方厘米)、 叶绿素( SPAD)、根长(厘米) 以 及鲜重和干重(克)。病原体拷贝数的减少通过 qPCR 定 量, 并根据之前在 2.4.1 节中描述的 TSWV-N 标准曲线进 行计算。 从每个重复中采集三株植物(每株植物取两片 叶), 标记并保存在 -20。C 下, 以测定病原体拷贝数的减 少量。利用以下公式计算内生菌处理对 TSWV 病原体的相 对减少率(%)。34
2.4.3 疾病严重程度评估及 AUDPC 计算
为在 0、7、 14 和 21 天后测定各处理中番茄斑萎病毒(TSWV) 的病情严重度(%), 采用 0 至 5 的分级标准, 其中 :0 = 无 症状; 1 = 幼叶出现轻微花叶;2 = 叶片出现花叶和皱缩;3 = 叶片出现黄斑并皱缩;4 = 叶片出现黄斑和坏死斑, 叶片变褐, 植株矮化;5 = 出现上述所有症状, 且植株部分萎蔫。35 数据 通过以下公式计算得出:
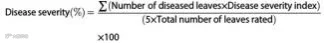
根据病情严重程度,按照建议在 0 至 21 天期间定期计算个体 治疗的累积病情发展评分(AUDPC)。
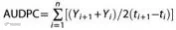
其中 Yi 表示疾病严重程度的比例,ti 表示时间(天数= 0、7、 14 和 21),n 表示观测总数。
根据病情严重程度和病情发展平均严重度(AUDPC)计算出 的对番茄斑萎病毒(TSWV)的生物防治效果,通过以下公式 得出:

2.4.4 酶激活反应的测定
该实验评估了内生菌对病毒感染植物中酶活性诱导的影响。
对包括丙二醛(MDA)、总酚、苯丙氨酸解氨酶(PAL)等 应激和防御相关指标以及过氧化氢酶(CAT)、多酚氧化酶 (PPO)、过氧化物酶(POD)、超氧化物歧化酶( SOD)等 抗氧化剂在内的几种酶进行了测定。此外,还按照第 2.1 节所 述的温室条件,对植物叶片中与防御相关的酶激活情况,如β- 1,3-葡聚糖酶和几丁质酶进行了测定。实验处理包括 DJB5、
L1-21、YN28、YN48、Mg6、I-CK 和 H-CK, 按照所述方法 采用喷雾法施用于植物。将每种处理中重复植物的叶片合并 为一个样本进行分析。
为提取并测定 CAT、PPO、POD 和 SOD 的活性, 将约 0.1 克 植物组织在冰浴中用 1 毫升提取缓冲液匀浆。混合物在 4℃、 12000 转/分钟的条件下离心 10 分钟, 上清液置于冰上保存。 CAT、PPO、POD 和 SOD 的抗氧化活性按照制造商(北京萨 拉生物科技有限公司)的方案进行测定。37
为提取和测定丙二醛(MDA),取约 0.1 克组织研磨成粉末, 加入 1 毫升提取液, 在冰浴中匀浆。 随后将样品在 4℃下以 8000×g 离心 10 分钟, 收集上清液并置于冰上直至检测。新 鲜样品约 0.1 克在 1.5 毫升 60%乙醇中匀浆, 测定总酚含量, 然后在 60℃下振荡 2 小时,接着在 25℃下离心 10 分钟。约 0.2 克植物组织用于测定苯丙氨酸解氨酶(PAL)活性, 将其在1 毫升提取缓冲液中于冰浴中匀浆。样品在 4℃下以 12000 转/分钟离心 10 分钟, 收集 上清液并置于冰上直至检测。按照制造商的说明,使用酶试剂 盒(北京索莱宝科技有限公司)测定 MDA、总酚和 PAL 活性。38
为测定β-1,3-葡聚糖酶和几丁质酶活性, 从处理过的植株 上采集 0.1 克新鲜离体叶片。将样本置于预冷的研钵中,加入 1 毫升 0.05 摩尔/升的乙酸钠缓冲液(pH 值 5.0)进行匀浆, 然后在 4℃下以 12000×g 离心 15 分钟。使用试剂盒(北京索 莱宝科技有限公司)按照制造商的说明测定β-1,3-葡聚糖酶和 几丁质酶的活性。39
2.4.5 统计分析
数据使用 Excel 软件 2016 版(美国华盛顿州雷德蒙德市微软 公司)进行分析, 并以均值 ± 标准误差的形式呈现。统计分 析使用 Statistix 软件( 8.1 版)和 Graph-Pad Prism 8( 8.0 版) 进行单因素和双因素方差分析(ANOVA)。 图表使用
GraphPad Prism( 8.0 版) 绘制 , 并使用 Adobe Illustrator CC 2019 进一步修改。统计设计包含四个生物学重复, 均值比较 采用 Tukey 多重范围检验, 显著性水平为 5%(P < 0.0001)。
3 结果
3.1 质粒构建和 qPCR 标准曲线
为了对番茄斑萎病毒(TSWV)进行定量检测,利用pMD-19 @ T 载体(Takara 公司)构建了一个 3320 个碱基对的 TSWV-N质粒, 并通过菌落 PCR 进行了验证, 得到了一条 628 个碱基 对的条带(支持信息, 图 S1(C))。随后, 对 PCR 产物进行 纯化、测序, 并与从 NCBI 数据库中检索到的序列进行比对, 以确认质粒中存在 TSWV-N 片段。此外, 还生成了用于绝对 定量 TSWV 的 qPCR 标准曲线, 其斜率为 -3.17,R2 = 0.988, 扩增 TSWV-N 的效率为 106.78%(图 S1(B))。
3.2 潜在细菌菌株的系统发育分析
如方法所述,从本实验室的培养库中总共获得了 15 株内生菌, 并通过半叶测定法对其对番茄斑萎病毒(TSWV)的生物防治 活性进行了筛选, 通过实时荧光定量 PCR(qPCR)对半叶中 的病原体拷贝数进行了定量。 其中, 包括解淀粉芽孢杆菌
DJB5、YN28、YN48 和 Mg6 以及枯草芽孢杆菌 L1-21 在内的 一些菌株对 TSWV 表现出显著的抑制作用, 因此被选作进一 步研究的对象(图 S1(C))。这些内生菌基于 gyrB 基因进行 PCR 扩增后进行了测序。所得的系统发育树表明,L1-21 与枯 草芽孢杆菌的亲缘关系很近(> 99.50%), 而 DJB5、YN28、 YN48 和 Mg6 则与解淀粉芽孢杆菌的同源性很高(> 99.50%), 且每株内生菌都处于不同的分支(图 S1(B))。
3.3 内生菌对番茄斑萎病毒的生物防治作用
在温室中进行了一项实验,以评估这些内生菌菌株抑制植物 体内番茄斑萎病毒(TSWV)感染的潜力。与对照处理相比, 不同内生菌(枯草芽孢杆菌 L1-21 和解淀粉芽孢杆菌 DJB5、 YN28、YN48 和 Mg6)处理在接种后不同天数(DPI)显著 (P < 0.0001)降低了本氏烟叶片中的 TSWV 拷贝数(图 1)。
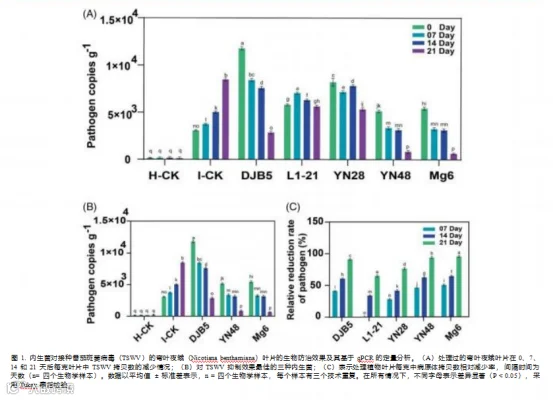
用解淀粉芽孢杆菌 DJB5 处理的植物, 其每克组织中番茄 斑萎病毒(TSWV) 的拷贝数从 0 天后(DPI) 的 1.18× 10^4 降至 7 天后的 8.43× 10^3, 减少了 28.5%, 相对生物防治效果 为 41. 18%。与 0 天后相比, 14 天后每克组织中的病毒拷贝数 降至 7.58× 10^3(减少了 35.7%, 相对生物防治效果为 60. 65%),21 天后降至 2.86× 10^3(减少了 75.7%, 相对生物防 治效果为 91.20%)。
用枯草芽孢杆菌 L1-21 处理的植株, 其病原体拷贝数从 0 DPI 时的 5.81× 103 呈上升趋势, 到 7 DPI 时达到 7.04× 103,病原 体载量减少了 21.2%, 相对生物防治效果为 0.35%。随后, 每 克植株的病原体拷贝数略有下降, 在第 14 天降至 6.30× 103 (减少 8.4%, 相对生物防治效果为 33.65%), 在第 21 天降 至 5.63× 103(减少 3.2%,相对生物防治效果为 64.91%)。
结果表明,枯草芽孢杆菌 YN28 在植株内对番茄斑萎病毒 (TSWV)具有生物防治效果, 在 7 天后(DPI)每克植株中 的病毒拷贝数从 8.21× 103 降至 7.17× 103, 使病原体减少了 12.7%, 相对生物防治效果为 28.21%。在 14 天后(DPI)(7. 82× 10 3)和 21 天后(DPI)(5.34× 10 3), 每克植株中的病 毒拷贝数略有下降, 病原体滴度分别降低了 4.8% 和 34.9%, 相对生物防治效果分别为 76.42% 和 28.09%。
内生菌枯草芽孢杆菌 YN48 在发病后 7 天显著降低了每克病原 体的拷贝数,从 5.14× 103 降至 3.35× 103,病原体滴度降低了 34.7%, 相对生物防治效果为 46.33%。随后, 在发病后 14 天和21 天, 每克病原体的拷贝数分别降至 3.14× 103 和 8.41× 102, 病原体滴度分别降低了 39.0% 和 83.6%, 相对生物防治效果 分别为 62.64% 和 90.06%。
同样,在 B. amyloliquefaciens Mg6 处理的植物中, 从 0 到 7天后 , 每克植物中 TSWV 的拷贝数从 5.43× 103 降至 3.26×103, 减少了 40.0%, 相对生物防治效果为 50.64%。在 14 天 和 21 天后, 由于内生菌的持续作用, 每克植物中的拷贝数分 别降至 3.13× 103 和 6.38× 102, 病原体的拷贝数分别减少了 42.3% 和 88.2%, 相对生物防治效果分别为 64.68% 和 95.73% (图 1(B)、(C))。
在 21 天后, 内生菌 DJB5、L1-21、YN28、YN48 和 Mg6
对番茄斑萎病毒的相对生物防治效果分别为 91.21%、64.92%、 76.43%、94.07% 和 95.73%, 与阴性对照组(I-CK)相比。 因 此, 与最初感染且病原体载量有所减少的阳性对照组和处理 组不同, 在任何时间间隔(7、 14 和 21 天后)均未在阴性对 照组中观察到病原体载量的明显减少, 如图 1(A)、(B)
所示。
3.4 病害严重程度与病情发展总值(AUDPC)
内生菌处理过的植物随时间推移病情严重程度的变化情况如图
2 所示。结果表明
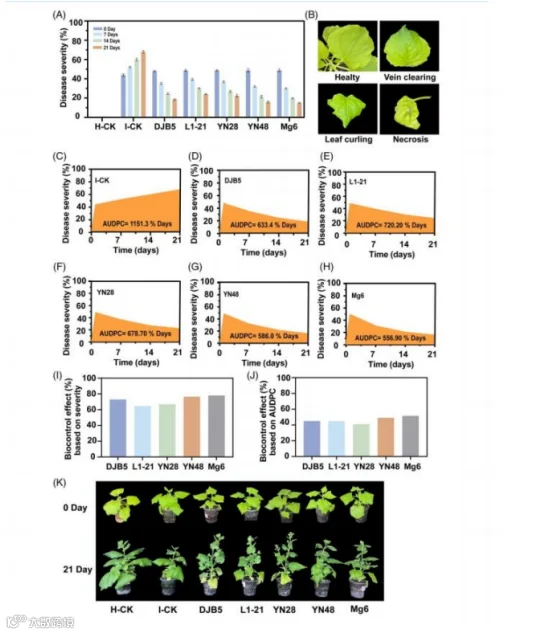
与对照组(I-CK)相比, 内生菌处理的植物中, 由内生菌掩 盖症状导致的病情严重程度呈下降趋势(图 2(B)、(K))。在 各处理组中,Mg6 和 DJB5 的病情严重程度最低, 分别为 28. 40% 和 31.55%。随着时间的推移,病情严重程度从 0 天后的 41.00% 下降到 21 天后的 23.40% 和 7 天后的 32.26%, 表明病 情严重程度随时间显著下降。 内生菌随时间推移的效力对病情 严重程度影响显著,Mg6 从 0 天的 48.80% 降至 21 天的 15%, YN48 从 0 天的 48.80% 降至 21 天的 16.00%(图 2(A))。
根据病情严重程度,内生菌 Mg6 和 DJB5 的最小累积病 情指数(AUDPC)值分别为 556.90% 和 633.4%。相比之下, L1-21 的最大 AUDPC 值为 720.20%, 表明其病情严重程度随 时间推移有所增加, 高于对照 I-CK( 1151.3%)。总体而言, 内生菌处理的植物 AUDPC 值随时间推移呈下降趋势,这表明 这些处理在减轻病情严重程度方面具有显著效果(图 2(C)- (H))。基于病情严重程度和 AUDPC 值, 内生菌 Mg6 对番茄 斑萎病毒(TSWV) 的总体生物防治效果分别为 78% 和 51. 65%, 其次是 YN48(分别为 76% 和 49. 10%)、DJB5(分别 为 73% 和 45%)、L1-21(分别为 64% 和 44.98%)和 YN28 (分别为 67% 和 41.04%), 与对照 I-CK 相比(图 2(I)、(J))。
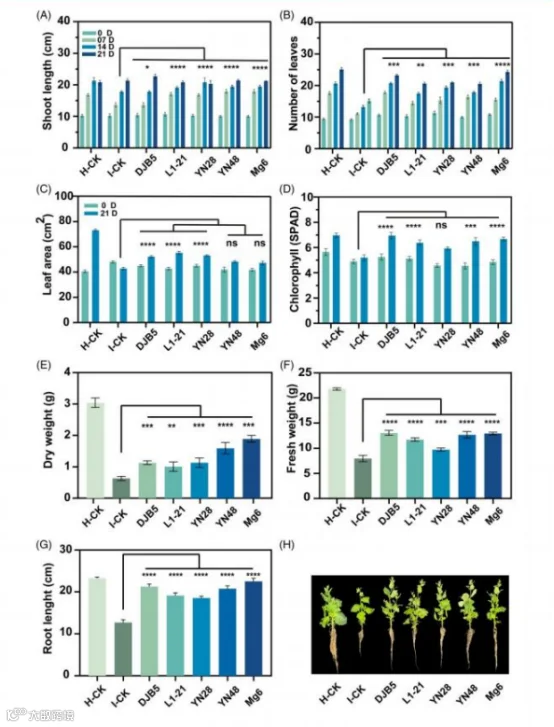
3.5 内生菌对植物生长的影响
植物生长参数在内生菌施用后显著(P < 0.0001)得到促进,
与对照处理相比,不同程度地减轻了番茄斑萎病毒的感染(图 3)。在所有处理中,YN48 的株高最高( 17.18 厘米), 其次 是 Mg6( 17.13 厘米), 与 I-CK 处理和其他内生菌处理相比。 在 21 天后,DJB5 和 YN28 的株高因时间间隔和内生菌处理的 相互作用而显著增加, 分别为 22.90 厘米和 21.38 厘米(图 3 (A))。在不同处理中,DJB5 和 Mg6 的叶片数量最多, 分别 为 18.12 片和 18.00 片, 高于 I-CK 处理。在时间间隔和处理 的相互作用下, 每株植物的叶片数量增多,DJB5 在 21 天后 有 23.25 片叶,YN28 有 20.98 片叶(图 3(B))。就叶面积而 言,YN28 的叶面积最大, 为 49.02 平方厘米, 其次是 L1-21 (48.95 平方厘米), 而 Mg6 的叶面积最小, 为 44.44 平方厘 米, 与 I-CK 处理无显著差异。 内生菌与时间间隔的相互作用 表明,L1-21 和 DJB6 的最大叶面积分别为 55.22 平方厘米 2 和 52.16 平方厘米 2, 与 I-CK 处理相比(图 3(C))。对于叶绿素 含量,DJB5 导致叶绿素含量最大增加(6.1 SPAD), 其次是 Mg6(5.76 SPAD), 与对照处理相比(图 3(D))。时间间隔 与内生菌的相互作用表明, 与 0 DPI 相比,21 DPI 时 DJB5 处 理的植物叶绿素含量最高(6.95 SPAD), 其次是 Mg6 处理的 植物(6.67 SPAD)。对于根长,Mg6 和 DJB5 内生菌分别比 I-CK 处理增加了 22.53 厘米和 21.25 厘米(图 3(G)、(H))。 用内生菌 DJB5 和 Mg6 处理的植物鲜重最高, 分别为 13.02 克和 12.93 克, 与 I- CK 处理相比(图 3(F))。关于干重,Mg6 和 YN48 处理的植 物干重最大,分别为 1.88 克和 1.59 克, 而对照处理的干重为 0.63 克(图 3(E))。
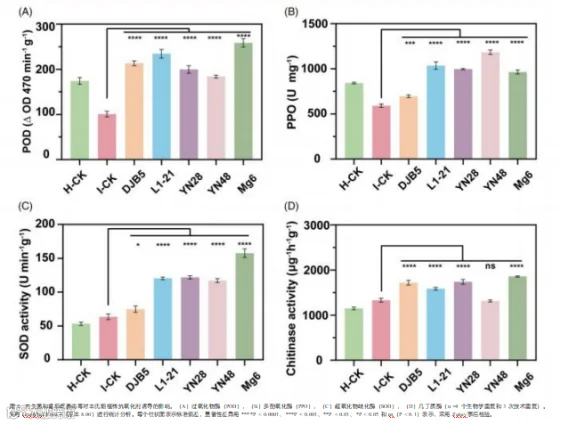
3.6 植物酶对内生菌和番茄斑萎病毒施用的反应
在 7 天后,对包括过氧化物酶(POD)、几丁质酶、多酚氧化 酶(PPO)和超氧化物歧化酶( SOD)在内的抗氧化酶的表达 进行了评估, 以探究内生菌处理对病毒感染植物氧化应激反应 的贡献以及对植物损伤的保护作用。与对照处理相比, 内生菌 处理显著(P < 0.0001)诱导了这些抗氧化剂的产生(图 4)。 内生菌的应用显著激活了植物的先天防御系统, 主要是通过诱 导酶活性。这种激活导致感染植物的病毒滴度显著降低, 并减 轻了病毒症状。因此,处理过的植物表现出更好的生长参数和 整体活力, 突出了内生菌在增强植物抗病性方面的作用(图 1、 2(A)、(K)和 3)。
在 H-CK 处理组中观察到过氧化物酶(POD)活性的最大值, 为 174 毫克/克, 而 I-CK 处理组的最小值为 101 毫克/克。接 种内生菌 Mg6、L1-21、DJB5、YN28 和 YN48 的植物中,
POD 活性显著提高 , 分别提高了 255.94%、232.47%、211. 09%、 197.53% 和 182. 18%, 均高于 I-CK 和 H-CK 处理组(图 4(A))。
I-CK 的超氧化物歧化酶( SOD)活性为 63.71 U min-1 g-1 , H-CK 的为 53.31 U min-1 g-1 。 内生菌处理的植物的 SOD 活 性高于对照处理,Mg6、YN28、L1-21、YN48 和 DJB5 处理 的植物的 SOD 活性分别比 I-CK 高 247.42%、 191.29%、 188. 99%、 183.77% 和 117.86%(图 4(C))。 同样, 内生菌处理的 植物在 H-CK 中表现出最高的多酚氧化酶(PPO)活性, 其值 为 842.7 U mg-1 , 在 I-CK 中观察到的最小值为 591.3 U mg- 1 。 与 I-CK 相比 , 内生菌 DJB5、L1-21、YN28、YN48 和 Mg6 分别使 PPO 活性提高了 117.68%、 175.21%、 168.70%、 200.55% 和 163.02%(图 4(B))。在几丁质酶活性方面 ,H- CK 和 I-CK 的值分别为 1151 和 1334 μg h-1 g-1 。在内生菌 处理下 ,Mg6( 139.23%)、YN28( 130.53%)、DJB5( 128. 95%)、L1-21( 118.98%)和 YN48(98.72%) 的几丁质酶活 性相比 I-CK 有最大幅度的增加。 同时,YN48 处理的植物的 几丁质酶活性无显著差异(图4(D))。
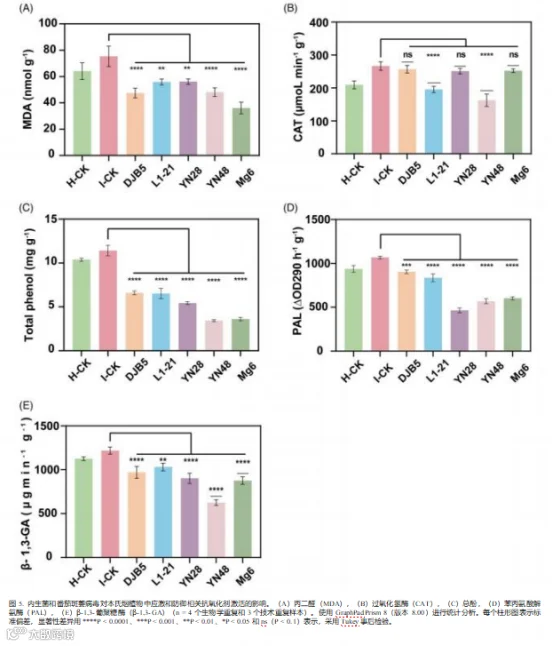
在 7 天后对 PAL(酶)、MDA(化合物)和总酚的水平 进行了分析, 以了解植物对病毒胁迫的反应及其防御机制。在 内生菌处理的植物中,PAL、MDA 和总酚的水平与对照处理 相比显著降低(P < 0.0001)(图 5)。实验中 ,I-CK 和 H- CK 的 PAL 值分别为 1065 和 937.3 (△OD290 h-1 g-1)。
PAL 活性的降低范围在最大和最小百分比值分别为 56.43%、 46.62%、43.54%、21.43% 和 15.06%。
图 2. 内生菌对感染番茄斑萎病毒(TSWV)的本氏烟叶片病害严重程度百分比和病情发展指数(AUDPC)的影响。(A)在接种 TSWV 和内生菌后 0、7、 14 和 21 天, 通过 0 至 5 的视觉评分标准评估植株的病害严重程度;(B)接种后植物叶片上出现的各种 TSWV 症状;(C-H) 内生菌处理的植株 AUDPC 值(即面积)从(D)到 (H)逐渐降低,表明内生菌在整个生长季内对 TSWV 题量有总体的抑制作用;(I,J)基于病害严重程度和 AUDPC 的生物防治效果;(K)接种 TSWV 后 0 天和 21 天内生菌对缓解植物 TSWV 症状的视觉效果。
结果表明,通过内生菌的应用来评估几丁质酶和β-1,3-葡聚糖酶的酶活性, 以确 定它们在植物抵御病原体中的作用。统计分析表明, 所有内生菌处理的植物与 对照处理相比, 差异显著(P
< 0.0001)(图 5)。从结果来看,H-CK 和 I-CK 的 CAT 值分别为 209.4 和 266.
8 μmoL min-1 g-1 , 其中 H-CK 的活性较高。 与 I-CK 相比,L-21 和 YN28 的 CAT 活性具有统计学意义。然而, 与 I-CK 相比,DJB5、L1-21、YN28、YN48 和 Mg6 的总体 CAT 活性分别降低了 3.84%、26.73%、5.83%、39. 11% 和 5.32% (图 5(B))。对于β-1,3-葡聚糖酶 ,H-CK 和 I-CK 的观测值分别为 1125 μg-1 min-1 g-1 和 1217 μg-1 min-1 g-1 。 内生菌处理的结果表明,YN48(48.61%)、 Mg6(28.01%)、YN28(25.88%)、DJB5(20.33%)和 L1-21(15.37%) 表现 出β-1,3-葡聚糖酶的激活, 与 I-CK 相比呈下降趋势(图 5(E))。
图 3. 内生菌对接种番茄斑萎病毒(TSWV)后本氏烟植株生长的影响。 内生菌促进了生长参数;(A)0、7、 14 和 21 天后植株的茎长(n =4 个生物样本),(B)0、7、 14 和 21 天后植株的叶片数量(n =4 个 生物样本),(C)0 和 21 天后植株的叶面积(n =4 个生物样本),(D)0 和 21 天后植株的叶绿素含量(n =4 个生物样本,每株植物测量 4 片叶),(E、F)21 天后植株的干重和鲜重(n =4 个生物样本), (G、H)21 天后植株的根长(n =4 个生物样本)。叶绿素含量以仪器读数的 SPAD 单位表示。SPAD 值的最大值表明叶片中叶绿素含量最高。 每个柱状图表示标准偏差, 显著性差异用 Tukey 事后检验表 示 :****P < 0.0001, ***P < 0.001, **P < 0.01, *P < 0.05,ns 表示 P < 0.1。
图 5. 内生菌和番茄斑萎病毒对本氏烟植物中应激和防御相关抗氧化剂激活的影响。(A)丙二醛(MDA),(B)过氧化氢酶(CAT),(C)总酚,(D)苯丙氨酸解 氨酶(PAL),(E)β-1,3-葡聚糖酶 (β-1,3-GA)(n = 4 个生物学重复和 3 个技术重复样本)。使用 GraphPad Prism 8(版本 8.00)进行统计分析。每个柱形图表示标 准偏差, 显著性差异用 ****P < 0.0001、***P < 0.001、**P < 0.01、*P < 0.05 和 ns(P < 0. 1)表示,采用 Tukey 事后检验。
4 讨论
内生菌是生活在植物内部的有益微生物,在正常和逆境条件下都能促进植物健康。40-42 此外, 这些微生物还能 抑制植物病害。43 自从发现有益微生物可作为天然生物 农药以来, 它们已成为控制植物病原体最有效的方法, 从而促进可持续农业 和全球农业安全。44 以往关于生物防治的研究大多集中 在细菌和真菌病害上, 而对于病毒的生物防治研究则相 对较少。对病毒进行系统且早期的检测是预防病毒传播 的最佳方法。45作为本研究中所展示的基于 qPCR 的方法通过将TSWV-N 克隆到 质粒中创建标准曲线, 对每克叶片中 TSWV-N 的拷贝数进行 了定量。标准曲线的斜率为 -3.17,R2 值为 0.988,效率为 106. 78%, 这表明 qPCR 的高效性和可靠性。qPCR 系统在检测范 围跨越多个数量级时更灵敏、更稳健, 可重复检测低至 1000 个 TSWV 目标分子。46
在本研究中,筛选出的内生菌 DJB5、YN48 和Mg6 对番 茄斑萎病毒(TSWV)表现出更强的拮抗活性, 与对照组(I- CK)相比, 能显著降低植物中的病毒滴度。具体而言, 在接 种后 21 天(DPI), 内生菌 Mg6(88.2%)、YN48(83.6%) 和 DJB5(75.7%)对病原体滴度的抑制效果最为显著(见图 2 (B)、(C))。Vitti 等人47 报道, 哈茨木霉 T22 菌株作为 一种生物防治剂, 可能是化学处理的替代品。它能通过激素信 号通路有效增强番茄植株对黄瓜花叶病毒(CMV)的抗性。
这种增强作用可能归因于内生菌在植物体内定殖,并通过生态位和营养竞争抑制病原微生物。先前的研究表明,微生物分泌的代谢产物48 能够通过直接限制病原体生长或间接创造不利环 境的方式抑制多种病原体。49,50
本研究揭示,内生菌能够减轻番茄斑萎病毒(TSWV)在 本氏烟(N. benthamiana)上的症状, 并通过降低病毒载量来 减轻病情。 同样 ,Beris 等人51 证明 , 解淀粉芽孢杆菌能将TSWV 病害的发生率降低 80%, 并延缓病毒积累。一些内生 菌分泌各种化合物52, 这些化合物能诱导植物产生生理变化,
从而减轻病害的可见症状,即使叶片中的病原体浓度很低也是 如此。我们的研究结果进一步支持了先前的发现,即芽孢杆菌、假单胞菌、链霉菌和其他物种在自然界中大量存在。53 这些物 种通过产生抗菌化合物、形成生物膜、从病原体中夺取营养以 及激活防御系统来保护寄主植物免受植物病原体的侵害。54 我 们的研究表明 , 解淀粉芽孢杆菌的分离株能够有效控制TSWV, 是一种可行的替代方案。通过将这些内生菌引入农业 实践, 我们可以有机地增强植物的防御能力, 减少对化学农药 的需求。这种方法为田间控制番茄斑萎病毒提供了一种实用且 环保的策略, 同时还有可能提高作物的产量和质量。
我们的研究表明,与对照组相比,这些内生菌有可能促进 植物生长参数,如叶片数量、茎和根的长度、叶面积、叶绿素 含量以及鲜重和干重(图 3)。大量研究已证实内生菌对植物 生长参数具有有益影响, 包括增加植物高度、生物量和整体活 力。55 内生菌通过提高氮和磷等养分的吸收以及促进赤霉素和 生长素等植物激素的生成来促进植物生长。56 我们的研究发现, 内生菌处理组的植物生长情况优于对照组,这支持了观察到的 生长改善情况。
近期的多项研究已表明,内生微生物能诱导植物产生相关 防御酶和抗氧化剂,如丙二醛(MDA)、苯丙氨酸解氨酶 (PAL)、总酚、多酚氧化酶(PPO)、过氧化物酶(POD)、 过氧化氢酶(CAT)、超氧化物歧化酶(SOD)、几丁质酶 和β-1,3-葡聚糖酶, 这可能有助于通过激活防御系统来保护植 物免受病原体侵害。57-60 我们的研究结果表明, 感染番茄斑萎 病毒(TSWV)的植物经内生菌处理后,通过显著提高多酚氧化酶(PPO)、过氧化物酶(POD)、超氧 化物歧化酶(SOD)和几丁质酶的活性,植物健康状况显著改 善, 这表明内生菌参与了植物防御相关酶的激活(图 4 和图 5)。在本研究中, 两种芽孢杆菌的应用均激活了植物中的酶 活性和应激相关酶, 从而降低了 TSWV 的感染。植物在应对 诸如真菌、细菌和病毒等植物病原体时产生应激和防御相关酶 及抗氧化剂的情况已有大量文献记载。61,62 关于多酚氧化酶 (PPO)、过氧化物酶(POD)、超氧化物歧化酶(SOD)和 几丁质酶的结果与Padro 等人的研究结果一致, 他们报道了用 枯草芽孢杆菌处理的番茄植株中抗氧化剂的显著增加。
在抗氧化酶中,超氧化物歧化酶(SOD)、过氧化物酶 (POD)、几丁质酶和多酚氧化酶(PPO)的协同作用对于显 著减轻氧化应激至关重要, 最终将细胞内的自由基维持在对植 物健康无害的水平。64 然而,抗氧化剂的水平取决于细菌克服 植物氧化应激的能力 。 已有研究对几种植物根际促生菌
(PGPR)进行了研究, 这些菌种能够维持植物细胞内的氧化 应激水平, 并促进植物生长。63,65 本研究结果表明, 与对照处 理相比, 感染植物经内生菌处理后, 丙二醛(MDA)、苯丙 氨酸解氨酶(PAL)、 总酚、β-1,3-葡聚糖酶和过氧化氢酶 (CAT)的含量均有所下降。这可能是由于内生菌与感染植物 相互作用, 诱导植物在病原胁迫下产生防御酶,如多酚氧化酶 (PPO)、过氧化物酶(POD)、超氧化物歧化酶(SOD)和 几丁质酶,通过清除过量的活性氧(ROS)来维持细胞内部的 完整性。丙二醛含量的降低表明内生菌在病原菌胁迫下有效减 少了膜的氧化损伤。
此外,在各种植物物种中观察到的氧化应激条件下 CAT 活性的降低可能与 SA 途径的激活有关, 从而导致 SA 的过量 释放。66,67 为了激活植物抵御病原体攻击的防御机制, 酶 PAL 对酚类化合物的合成至关重要。68 内生菌处理 7 天后,植物体 内酚类化合物和 PAL 的浓度降低表明内生菌通过显著控制植 物内的感染发挥了生物防治作用。最初,在感染期间酚类化合 物的浓度会增加。Kobyletska 等人报告了防御酶浓度降低的趋 势,69 其中植物在施用外源 SA 后会控制感染以对抗病原体。 对于参与病原体降解的 囚-1,3-葡聚糖酶也观察到了类似的趋 势。结果表明, 随着内生菌处理 7 天后病毒载量的降低,植物 体内 囚-1,3-葡聚糖酶的水平也降低, 以维持内部环境。
5 结论
总之,我们的研究证明,诸如枯草芽孢杆菌 DJB5、YN48 和 Mg6 等内生菌能够有效控制番茄斑萎病毒(TSWV)对本氏烟 的感染, 在 21 天后持续减轻症状, 降低病情严重程度, 并降 低累积病情指数(AUDPC)。这些内生菌还显著改善了生长 参数, 增加了茎和根的长度、叶片数量、叶面积和生物量。此 外, 内生菌的应用激活了关键的抗氧化酶(多酚氧化酶、过氧 化物酶、超氧化物歧化酶和几丁质酶), 同时降低了应激和防 御相关指标(丙二醛、苯丙氨酸解氨酶、总酚和β-1,3-葡聚糖 酶),从而保护植物免受病毒引起的损害。 由于番茄斑萎病毒 在茄科作物中造成了重大损失, 因此迫切需要探索基于自然的 解决方案。我们的研究强调了内生菌作为生物防治剂提高作物 产量的潜力,并为番茄斑萎病毒的管理提供了新的见解。
作为 IDM 的一部分, 对特定植物病毒及其他植物病毒进 行研究。有必要进一步探究内生菌诱导的防御反应的分 子机制, 这将为优化和扩大其在可持续农业中的应用提供宝贵见解。(本文为AI中文译文,如有错谬为整理者的失误)
(参考文献略)



1.速效:本靶标生防菌株能够迅速阻止(溶解)病毒蛋白合成,作物迅速恢复健康。
2.长效:本内生菌在植物体内长期持续扩繁,长期发挥防控作用,预防病病毒发生。
3.活性:本内生菌在植物体内定殖后不断扩繁增量,持续发挥防控作用。
4.安全:产品通过有机投入品认证,对植物、动物、昆虫、环境和人类等安全友好。
5.抗逆:改善植株微生态环境,诱导作物产生免疫力,增强抗逆性,促进作物生长发育。


1、本菌株为作物病毒的靶标内生菌,通过叶片或根系进入作物体内后迅速定殖扩繁,分泌的活性物质对各种病毒迅速产生高抗性,阻止(溶解)病毒蛋白合成,迅速降低病毒数量,作物新叶新梢迅速恢复正常。防治效果达到95.7%。
2、本菌株进入植物体内后,迅速改善植株体内微生态环境,提高作物免疫力,抗逆性显著增强,减轻病毒对作物的危害。显著促进作物生长发育,增加茎和根的生长、叶片数量、叶面积和生物量等。
3、本菌株能够增强作物光合作用,叶片浓绿油亮,健壮植株,促进作物恢复健康。作物发病时加大浓度施用,效果更为显著。



黄瓜使用图片:



云南省石屏县杨梅使用图片:杨梅双生病毒
未喷洒丙度七号前病状:

喷洒丙度七号之后表现:



烟草使用图片:


榨菜使用图片:

视频反馈效果:(扫面二维码可了解使用效果)

*注意事项:
1、本品使用时不得与化学杀菌剂接触,与上次喷洒杀菌剂要有间隔期,国产杀菌剂不低于5天,进口杀菌剂不低于7天。
2、本品为内生菌,必须喷透喷匀,建议连续喷洒三次以上。作物不健康状态时高浓度喷洒1-2次,然后再正常喷洒防控。作物正常防控参照使用说明。
3、建议在傍晚或阴天喷洒,大棚阴雨天可全天喷洒,菌液在叶片湿润时间越长效果越好,尽量配用没有杀菌作用的助剂。
4、本品使用浓度越大效果越显著,首次使用建议三天内连喷两次,常规防控间隔7-10天连续使用。

权威研制,值得信赖!!
18164480171(史经理)