

1.1 核孔复合体(NPC)
核质转运的核心是核孔复合体(Nuclear pore complex, NPC),这是嵌入细胞核膜上的一个大型蛋白质复合体,是控制物质进出细胞核的通道,参与大分子、核糖体亚基、病毒蛋白和RNA(mRNA、rRNA、tRNA、miRNA)的双向核-质运输。
NPC由大约30多个核孔蛋白 (nucleoporins, Nups) 组成,不同的核孔蛋白复合物以特定的排列方式构成了核孔复合体的结构,从细胞质排列到核质,构成细胞质丝、细胞质环(CR)、内孔环(IR)、核环(NR)和核篮(NB)。NPC内部存在富含苯丙氨酸 (Phe) 和甘氨酸(Gly) 的重复序列的Nup,称为FG-Nups,它们与核转运蛋白结合,从而促进核双向转运[1]。
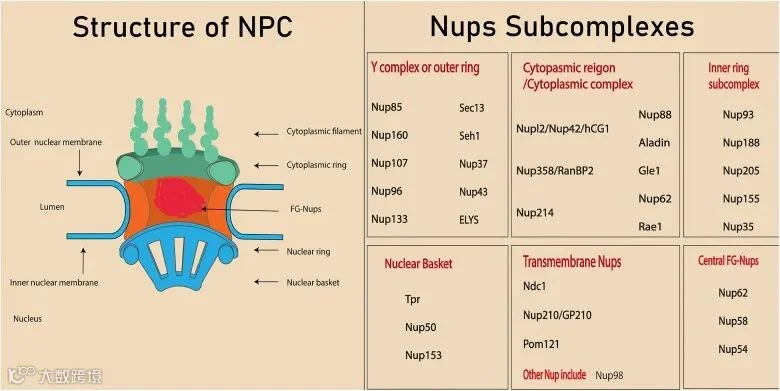
图1:核孔复合体的结构(图源:参考文献1)
1.2 核定位信号(NLS)与核输出信号(NES)
核质转运过程中,信号序列在确定分子进出细胞核的方向性上扮演着关键角色。小分子物质可以通过自由扩散或被动扩散穿过NPC通道,分子量大于40 kDa的蛋白质必须通过主动运输才能进出细胞核。因此,这些蛋白需要有核定位信号(nuclear localization signal, NLS)和核输出信号(nuclear export signal, NES)。
核定位信号(NLS)是一段特定的氨基酸序列,通常包含正电荷较多的赖氨酸和精氨酸残基,能够引导蛋白质进入细胞核。相对应的,核输出信号(NES)则是一段富含亮氨酸或其他疏水性氨基酸的序列,能够引导蛋白质从核内运出至核外。
NLS:典型的NLS是短而连续的氨基酸序列,如经典的SV40大T抗原中的 NLS,序列为PKKKRKV[2]。NLS通常位于蛋白质的暴露表面,便于核转运受体(如 Importin α)识别。
NES:通常是短而连续的氨基酸序列,但与NLS不同,NES的核心特征是富含疏水性氨基酸(如亮氨酸)。NES的共识序列为Φ1-X(2-3)-Φ2-X(2-3)-Φ3-X-Φ4基序(Φ:代表疏水残基:L、I、F、M或V;X:任何氨基酸)。后来,根据这些Φ的模式和间距序列,又将NES基序分为1a、1b、1c、1d、2、3和4类[3,4]。一个经典的NES序列是HIV Rev蛋白中的NES,序列为LQLPPLERLTL[5]。

图2:NES共识序列(图源:参考文献3)
这些信号序列使得蛋白质能够被识别并结合到相应的核转运受体上,从而确保蛋白质在核质之间的准确运输。
1.3 核转运受体(NTR)
核转运受体(Nuclear Transport Receptor, NTR)在核质转运过程中起着至关重要的作用,负责识别和结合带有NLS或NES的货物蛋白,并将其引导通过NPC。目前发现的NTR都属于核转运蛋白(karyopherin,KAP或KPN)家族。
按照结构和作用特点,核转运蛋白分为Karyopherin α(KPNA)和Karyopherin β(KPNB)两类。按照功能,核转运蛋白又可分为核输入蛋白(Importin,Imp)和核输出蛋白(exportin)。KPNA均为核输入蛋白,而KPNB则包括核输入蛋白和和核输出蛋白。
核输入蛋白(Importin,Imp):包括Importin α 和 Importin β 等,它们主要负责识别和转运携带NLS的蛋白质。Importin α 通常与携带NLS的蛋白质直接结合,但没有穿梭功能,需要由Importin β携带入核。Importin β具有穿梭核质的作用, 通过与Importin α形成复合物,将整个复合物通过NPC运入细胞核。
核输出蛋白(exportin):Exportins如核输出蛋白1(XPO1或CRM1)等,主要负责识别和转运携带NES的蛋白质。Exportins 通常通过结合Ran-GTP与NES一起形成复合物,并将其转运出细胞核。
人类的Importin 13和exportin 4都具有双向运输能力。
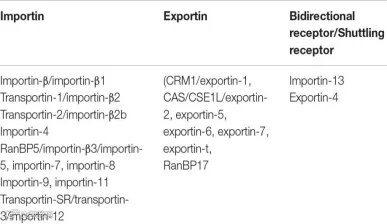
图3:核转运蛋白分类(图源:参考文献6)
核转运受体不仅负责核质转运的具体执行,还参与核质转运的选择性调控。例如,某些蛋白质只有在特定信号条件下才会暴露其NLS或NES,从而触发特定的核质转运过程。
1.4 小GTP酶Ran
核质转运另一个关键分子是小GTP酶Ran。与其他小GTP酶一样,Ran的功能通过与GTP或GDP结合来调节。Ran主要在细胞核中与GTP结合,在细胞质中与GDP结合。由于Ran-GEF在细胞核中催化Ran-GDP变为Ran-GTP,而Ran-GAP则在细胞质中催化 Ran-GTP水解为Ran-GDP,形成了一个Ran GTPase循环。
RanGTP与核转运受体复合物一起运输到细胞质。在细胞质中,RanBP1或RanBP2 (Nup358)促进RanGTP与核转运受体复合物的解离,允许RanGAP1介导的GTP水解。RanGDP通过NTF2循环回到细胞核,与NTF2解离,并通过染色质结合的RCC1 (GEF)重新装载GTP。除了这些必需因子外,四个Ran结合蛋白(RanBP1、RanBP2、RanBP3和Nup50)调节RanGTP与核细胞蛋白的相互作用,加快运输速度。

图4:RanGTP系统及调控蛋白(图源:参考文献7)
通常直径小于5 nm或分子量小于40 kDa的分子可以自由穿过NPC通道,而较大的分子就需要主动转运。
核输入:核转运受体(NTR)识别货物蛋白中特定的核定位序列(NLS)。例如,含经典核信号序列的货物蛋白与输入蛋白Importin-α结合,然后再与Importin-β形成三聚体复合物。如果NLS是非典型核信号序列,那么输入蛋白β直接与它的货物分子结合,而不需要输入蛋白α的参与。Importin α能够识别经典NLS,但没有穿梭功能,Importin β具有穿梭核质的作用,需要由Importin β携带入核。三聚体复合物由Importin-β1携带着通过NPC,进入细胞核,入核之后复合物解离,释放货物蛋白,货物蛋白行使其功能。
核输出:核输出是通过核输出序列(NES)的识别来实现的。核输出开始于Ran-GTP与核输出蛋白exportin 1(CRM1,XPO1)结合,这导致核输出货物的亲和力增加。然后,复合物移动到核孔中,由RCC1激活Ran-GTP水解,形成核输出复合物。复合物穿过NPC,在细胞质中,复合物解离,释放货物蛋白。
通过Ran“再循环”过程,使两种核转运蛋白返回细胞质,以进行下一轮转运。Importin-β1可以与Ran-GTP结合,通过NPC回到胞质;Importin-α因为缺乏穿梭能力,所以除需要Ran-GTP外还需要一个输出蛋白,如CAS(exportin2)或CRM1(exportin1)。

图5:核质转运机制信号通路(图源:参考文献8)
3.1 核质转运与神经退行性疾病
近年来,核质转运与神经退行性疾病之间的关系成为研究热点,特别是肌萎缩性侧索硬化症(ALS)、阿尔茨海默病(AD)和亨廷顿病(HD)。
越来越多的证据表明,核质转运缺陷导致某些关键蛋白质无法正确定位是ALS病理的关键机制。例如,散发性ALS脊髓运动神经元中核孔蛋白Nup62和核输入蛋白KPNB1(即Importin β1)核染色被破坏,TDP-43明显定位错误[9]。profilin1突变会导致家族性ALS 运动神经元的核质转运受损。在具有突变profilin1的运动神经元中,Ran、RanBP2和 RanGAP1在细胞质中分布错误[10]。
核质转运缺陷也是HD的一个致病因素,突变亨廷顿蛋白(mHTT)可能通过干扰核孔复合物、核孔蛋白和其他运输因子来破坏核质转运[11]。例如,使用小鼠模型、果蝇模型、人类患者来源的神经元和转染了全长mHTT的原代神经元,证明了HD基因内CAG三联体重复扩增的产物会导致核孔复合体和核质运输中断。在HD和JHD患者的死后脑组织中检测到RanGAP1和Nup62的错误定位和/或聚集,Nup62在几种HD动物模型、HD人类组织和HD ipsc衍生的神经元中定位错误和/或聚集[12]。
核孔形态和功能缺陷与阿尔茨海默病(AD)有关。tau蛋白是AD疾病进展相关标志物,tau蛋白可以直接与核孔复合体 (NPC) 的核孔蛋白相互作用并影响其结构和功能完整性[13,14]。在阿尔茨海默病(AD)及tau蛋白转基因小鼠模型中,核孔蛋白Nup98表现出显著的亚细胞定位异常,这一现象可能与AD的病理进程密切相关。另外,NRF2在AD发病机制中扮演着重要角色,在正常情况下NRF2的核转运由Importin-α5和β1介导,然而在AD患者中,NRF2的核转运过程明显受阻,导致其在细胞质中异常聚集[15]。
核质转运缺陷正在成为多种神经退行性疾病的重要病理机制,提示核质转运的调控可能是治疗这些疾病的新靶点。
3.2 核质转运与癌症
在癌症研究中,核质转运的异常被认为与肿瘤的发生和发展有密切关联。核孔蛋白Nup98也是白血病中已明确鉴定的原癌基因,也是在急性髓系白血病患者中观察到的复发性染色体易位事件的一部分[16]。核孔蛋白Nup88在卵巢癌、淋巴瘤、间皮瘤、多种肉瘤和一些上皮癌中过度表达,乳腺癌、结直肠癌和肝细胞癌中Nup88过度表达与肿瘤侵袭性有关[17]。
核输出蛋白1(XPO1)在骨髓瘤、淋巴瘤、卵巢癌、脑胶质母细胞瘤、骨肉瘤、胰腺癌、宫颈癌和胃癌等多种恶性肿瘤中过表达[18],XPO1可作为肿瘤患者预后判断的指标,也是抗肿瘤治疗的潜在靶标。
Ran是一种核质穿梭蛋白,有研究发现Ran是卵巢癌转移的“帮凶”,在卵巢癌细胞中,RhoA在Ran蛋白的帮助下才能够到达细胞膜,而RhoA在细胞迁移中起重要作用[19]。抑制Ran很可能是治疗卵巢癌以及其他癌症的有效策略。
核质转运在癌症的发生、发展、转移等过程中起着重要作用,针对核质转运相关蛋白或通路的研究可能为癌症的诊断和治疗提供新的靶点和策略。
3.3 核质转运与药物开发
在药物研发方面,针对蛋白质核质转运的药物设计具有广阔的前景。目前已经有一些药物在临床试验中显示出了潜在的治疗效果。
核输出蛋白1(XPO1)在肿瘤细胞中常过量表达,导致多种抑癌基因蛋白及生长调节蛋白亚细胞定位及功能紊乱,从而促进肿瘤生长与存活。XPO1抑制剂selinexor(塞利尼索),已被用于治疗复发难治性多发性骨髓瘤[20]和弥漫大B细胞淋巴瘤[21],为癌症治疗提供了新的方向和选择。
核输入蛋白,如Importin-β1、Importin-α2和Importin-α4,已被证明与不同肿瘤的发生和发展有关,针对Importin等核转运蛋白的抑制剂被认为是抗肿瘤治疗的潜在靶点。Importin-β特异性抑制剂importazole(IPZ)可阻断Importin-β介导的核输入,而不会破坏转运蛋白介导的核输入或XPO1介导的核输出[22]。相比于核输出蛋白抑制剂的快速发展,核输入蛋白特异性抑制剂的开发仍处于起步阶段。
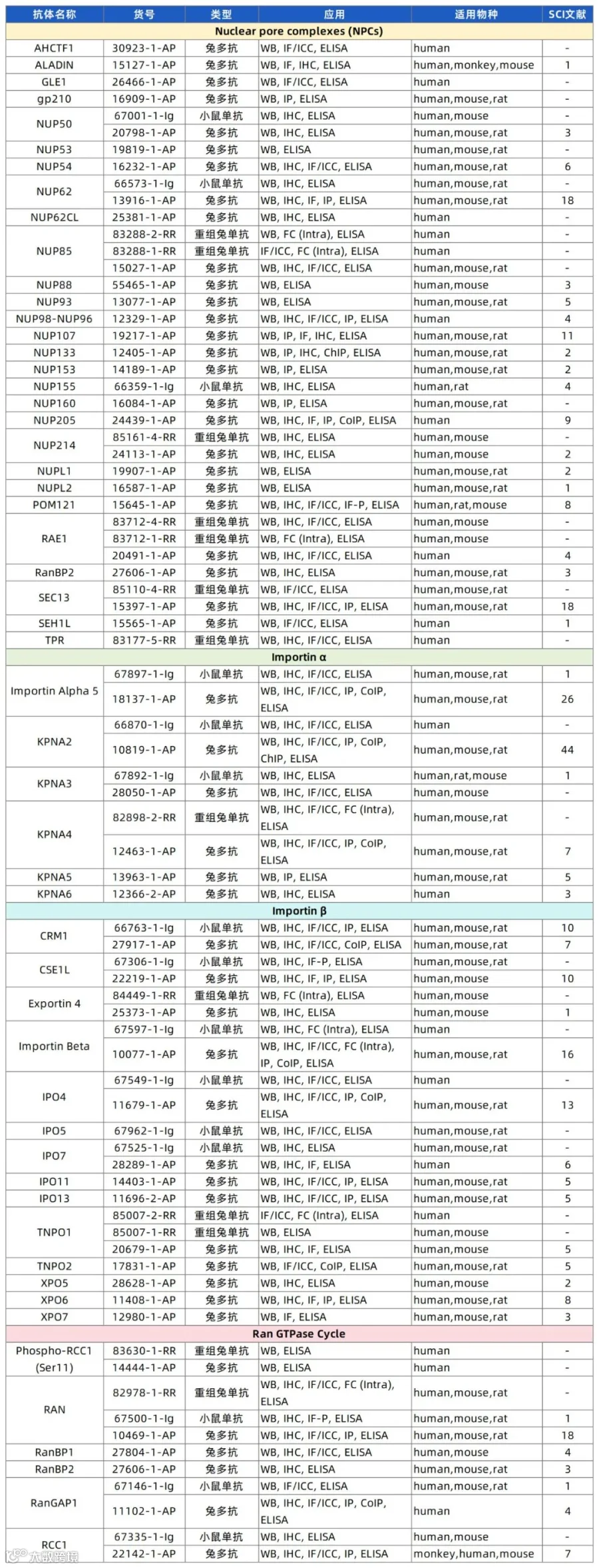
作为科研工作者最可靠的伙伴之一,Proteintech深耕抗体领域二十余年,产品覆盖生命科学领域。下面小P将核质转运相关抗体推荐给大家,希望对相关研究带来助力。
(上下滑动查看产品列表)
参考文献:
1. Khan AU, Qu R, Ouyang J, Dai J. Role of Nucleoporins and Transport Receptors in Cell Differentiation. Front Physiol. 2020 Apr 3;11:239.
2. Dingwall C, Laskey RA. Nuclear targeting sequences--a consensus? Trends Biochem Sci. 1991 Dec;16(12):478-81.
3. Lee Y, Pei J, Baumhardt JM, et al. Structural prerequisites for CRM1-dependent nuclear export signaling peptides: accessibility, adapting conformation, and the stability at the binding site. Sci Rep. 2019 Apr 29;9(1):6627.
4. Xu D, Farmer A, Collett G, et al. Sequence and structural analyses of nuclear export signals in the NESdb database. Mol Biol Cell. 2012 Sep;23(18):3677-93.
5. Fischer U, Huber J, Boelens WC, et al. The HIV-1 Rev activation domain is a nuclear export signal that accesses an export pathway used by specific cellular RNAs. Cell. 1995 Aug 11;82(3):475-83.
6. Oka M, Yoneda Y. Importin α: functions as a nuclear transport factor and beyond. Proc Jpn Acad Ser B Phys Biol Sci. 2018;94(7):259-274.
7. Yang Y, Guo L, Chen L, et al. Nuclear transport proteins: structure, function, and disease relevance. Signal Transduct Target Ther. 2023 Nov 10;8(1):425.
8. De Jesús-González LA, Palacios-Rápalo S, Reyes-Ruiz JM, et al. The Nuclear Pore Complex Is a Key Target of Viral Proteases to Promote Viral Replication. Viruses. 2021 Apr 19;13(4):706.
9. Aizawa, H.; Yamashita, T.; Kato, H.; et al. Impaired nucleoporins are present in sporadic amyotrophic lateral sclerosis motor neurons that exhibit mislocalization of the 43-KDa TAR DNA-binding protein. J. Clin. Neurol. 2019, 15, 62–67.
10. Giampetruzzi, A.; Danielson, E.W.; Gumina, V.; et al. Modulation of actin polymerization affects nucleocytoplasmic transport in multiple forms of amyotrophic lateral sclerosis. Nat. Commun. 2019, 10, 3827.
11. Ding B, Sepehrimanesh M. Nucleocytoplasmic Transport: Regulatory Mechanisms and the Implications in Neurodegeneration. Int J Mol Sci. 2021 Apr 17;22(8):4165.
12. Grima JC, Daigle JG, Arbez N, et al. Mutant Huntingtin Disrupts the Nuclear Pore Complex. Neuron. 2017 Apr 5;94(1):93-107.e6.
13. Eftekharzadeh B, Daigle JG, Kapinos LE, et al. Tau Protein Disrupts Nucleocytoplasmic Transport in Alzheimer's Disease. Neuron. 2018 Sep 5;99(5):925-940.e7.
14. Nag N, Tripathi T. Tau-FG-nucleoporin98 interaction and impaired nucleocytoplasmic transport in Alzheimer's disease. Brief Funct Genomics. 2023 Apr 13;22(2):161-167.
15. Ramsey, C.P.; Glass, C.A.; Montgomery, M.B.; et al. Expression of Nrf2 in neurodegenerative diseases. J. Neuropathol. Exp. Neurol. 2007, 66, 75–85.
16. Nakamura T, Largaespada DA, Lee MP, et al. Fusion of the nucleoporin gene NUP98 to HOXA9 by the chromosome translocation t(7;11)(p15;p15) in human myeloid leukaemia. Nat Genet. 1996 Feb;12(2):154-8.
17. Hill R, Cautain B, de Pedro N, Link W. Targeting nucleocytoplasmic transport in cancer therapy. Oncotarget. 2014 Jan 15;5(1):11-28.
18. Lai C, Xu L, Dai S. The nuclear export protein exportin-1 in solid malignant tumours: From biology to clinical trials. Clin Transl Med. 2024 May;14(5):e1684.
19. Zaoui K, Boudhraa Z, Khalifé P, et al. Ran promotes membrane targeting and stabilization of RhoA to orchestrate ovarian cancer cell invasion. Nat Commun. 2019 Jun 17;10(1):2666.
20. Mo CC, Yee AJ, Midha S, et al. Selinexor: Targeting a novel pathway in multiple myeloma. EJHaem. 2023 May 15;4(3):792-810.
21. Schuster M, Zijlstra J, Casasnovas RO, et al. Effect of Prior Therapy and Disease Refractoriness on the Efficacy and Safety of Oral Selinexor in Patients with Diffuse Large B-cell Lymphoma (DLBCL): A Post-hoc Analysis of the SADAL Study. Clin Lymphoma Myeloma Leuk. 2022 Jul;22(7):483-494.
22. Soderholm JF, Bird SL, Kalab P, et al. Importazole, a small molecule inhibitor of the transport receptor importin-β. ACS Chem Biol. 2011 Jul 15;6(7):700-8.
▲上下滑动查看更多


扫码添加云南总代理云南泽浩客服微信






往期文章推荐
