


摘要:本研究旨在探究酵母培养物(YC)对奶牛生产性能、血清生化指标、瘤胃微生物及代谢物的影响。选用泌乳天数、胎次、产奶量相近的荷斯坦奶牛40头,随机分为2组,对照组饲喂基础饲粮(不含YC),试验组饲喂基础饲粮+YC(每头每天100g),试验期35d,其中预试期7d,正试期28d。结果表明:与对照组相比,1)添加YC显著提高了奶牛的产奶量、乳脂产量、乳糖产量、乳蛋白产量(P<0.05),显著降低了体细胞数(P<0.05);2)添加YC极显著提高了血清中总蛋白含量(P<0.01),显著提高血清中白蛋白、钙、总胆固醇及低密度脂蛋白含量(P<0.05);3)添加YC显著提高了血清中过氧化氢酶和超氧化物歧化酶的活性(P<0.05);4)添加YC极显著增加了瘤胃微生物蛋白的含量(P<0.01),显著提高了氨态氮、乙酸、异丁酸、异戊酸、戊酸的含量(P<0.05),瘤胃总挥发性脂肪酸含量也有升高的趋势(P=0.051);5)添加YC显著提高了奶牛瘤胃中UCG-002、毛螺菌科UCG-004、未分类硒单胞菌科、未分类WCHB1-41等的相对丰度(P<0.05);瘤胃代谢组学共鉴定出224个差异代谢物,其中155个上调,69个下调。差异代谢物富集于维生素B6代谢、初级胆汁酸生物合成、色氨酸代谢和α-亚麻酸代谢4个通路。综上所述,饲粮中添加YC可以提高奶牛瘤胃特定细菌丰度,增加挥发性脂肪酸、微生物蛋白和B族维生素等合成,改善瘤胃发酵效率并增强奶牛抗氧化能力,进而提高泌乳后期奶牛的生产性能。
关键词:酵母培养物;奶牛;生产性能;瘤胃微生物;瘤胃代谢物
酵母培养物(YC)是将酵母细胞在特定的培养条件下,通过特定发酵工艺形成的一种酵母发酵产品,除含有酵母细胞内的营养物质外,还含有丰富的酵母代谢产物,如小肽、甘露寡糖、β-葡聚糖、氨基酸、未知生长因子等,营养较为全面。研究表明,饲喂YC可以提高奶牛对饲粮的消化率,提高生产性能、抗氧化能力,改善瘤胃发酵,维持瘤胃稳态。Halfen等研究发现,YC可以增加泌乳中期奶牛瘤胃中纤维分解菌和乳酸分解菌的占比,降低淀粉分解菌的占比,提高奶牛对饲料的利用率。Desnoyers等研究表明,饲喂YC可以改善奶牛的生产性能、稳定瘤胃pH、增加挥发性脂肪酸含量和乙丙比。Dias等报道,添加YC可以提高奶牛的泌乳性能,降低奶牛亚急性瘤胃酸中毒的风险。奶牛在泌乳后期采食量下降,饲粮中粗饲料的比例升高,但其营养需求仍然较高,添加YC可以改善瘤胃功能,提高能量利用率。此外,YC还具有增强奶牛免疫功能和缓解氧化应激的作用,可以改善奶牛机体健康。且现有文献大多关注YC在奶牛泌乳盛期的作用,很少有对泌乳后期奶牛添加YC的效果及其作用机理的报道。因此,本试验通过在饲粮中添加YC,探究其对泌乳后期奶牛的影响,从瘤胃微生物的角度出发,通过16SrRNA高通量测序技术与瘤胃液非靶向代谢组学联合分析来进一步阐明YC的作用机理,为YC在生产中的使用提供理论依据。
YC蛋白质含量>16%,甘露聚糖含量≥3%,水分含量<12%。
本次动物试验已由西北农林科技大学实验动物管理及伦理审查执行委员会批准,审批号:NWAFU-DK2024016。
本试验在宁夏某牧场进行。选用泌乳天数[(218±10)d]、胎次(3.5±0.5)、产奶量[(46±5)kg/d]相近的荷斯坦奶牛40头,随机分为2组,每组各20头,对照组(CON组)饲喂基础饲粮(不含YC),试验组(YC组)饲喂基础饲粮+YC(每天每头100g)。试验期35d,其中预试期7d,正试期28d。试验牛均采用散栏式饲养,每天投料2次(10:40、18:40),挤奶3次(06:00、14:00、22:00),自由饮水。基础饲粮组成及营养水平见表1。
表1 基础饲粮组成及营养水平(干物质基础)
项目Items |
含量Content |
原料Ingredients |
|
青贮玉米Cornsilage |
36.35 |
压片玉米Flakedcorn |
3.64 |
全棉籽Wholecottonseed |
5.54 |
苜蓿干草Alfalfahay |
3.27 |
苜蓿青贮Alfalfasilage |
6.70 |
玉米Corn |
16.02 |
豆粕Soybeanmeal |
12.46 |
棉籽粕Cottonseedmeal |
2.23 |
菜籽粕Rapeseedmeal |
2.45 |
项目Items |
含量Content |
麦麸Wheatbran |
1.56 |
碳酸氢钠NaHCO3 |
1.02 |
脂肪酸钙Fattyacidcalcium |
0.98 |
湿玉米酒糟Wetcorndistiller’sgrains |
4.45 |
膨化大豆Extrudedsoybean |
1.33 |
预混料Premix1) |
2.00 |
合计Total |
100.00 |
营养水平Nutrientlevels2) |
|
粗蛋白质CP |
17.13 |
粗脂肪EE |
5.56 |
中性洗涤纤维NDF |
31.36 |
酸性洗涤纤维ADF |
20.98 |
粗灰分Ash |
7.59 |
钙Ca |
0.94 |
磷P |
0.39 |
泌乳净能NEL/(MJ/kg) |
6.53 |
1)预混料为每千克饲粮提供The premix provided the following per kg of the diet:VA 100 000 IU,VD320 000 IU,VE≥1000 mg,Cu 300 mg,Zn 1000mg,Mn 600mg,I 10mg,Se 5mg,Co 5mg。
2)泌乳净能为计算值,其余为实测值。NEL was a cal-culated value,while the others were measured values.
粗蛋白质(CP)、粗脂肪(EE)和粗灰分(Ash)含量分别参照GB/T6432—2018、GB/T6433—2006、GB/T6438—2007方法测定;中性洗涤纤维(NDF)、酸性洗涤纤维(ADF)含量参考Van Soest等方法测定;钙(Ca)含量参照GB/T6436—2018方法测定;磷(P)含量参考GB/T6437—2018方法测定;泌乳净能(NEL)参照NRC(2001)计算。
试验期间每天记录2组的饲粮投喂量和剩料量。同时,采用四分法采集饲粮样,测定干物质含量,用每组实际采食量再除以每组的牛头数,计算各组的DMI。通过牧场利拉伐帝波罗挤奶系统记录试验牛只每天的产奶量,并于试验期的第7、14、21、28天采集早、中、晚各50mL乳样,按4:3:3的比例混匀后保存。送往吴忠市奶业研究院进行乳蛋白、乳脂、乳糖、非脂乳固体、总固形物、乳尿素氮及体细胞数的测定,并计算4%乳脂校正乳(FCM)产量和泌乳效率(FE),计算公式如下:4%FCM产量=0.4×产奶量(kg/d)+15×乳脂产量(kg/d);FE=4%FCM产量/DMI。
于试验期第28天,晨饲前用真空采血管采集牛尾静脉血液30mL,静置30min后,3000×g、4℃离心15min制备血清样品,分装于1.0mL离心管,-20℃保存待测。使用全自动生化仪(7020型,日立,日本)检测血清中总蛋白、白蛋白、总胆固醇、低密度脂蛋白、甘油三酯、葡萄糖、钙、肌酐含量。血清过氧化氢酶(CAT)、谷胱甘肽过氧化物酶(GHS-Px)、谷胱甘肽-S转移酶(GST)、超氧化物歧化酶(SOD)活性和丙二醛(MDA)的含量用试剂盒测定,试剂盒购自南京建成生物工程研究所。
于试验期28天,2组各随机选取8头奶牛,晨饲前使用瘤胃液口腔采集器采集奶牛瘤胃液样品,弃掉前50mL瘤胃液后采集100mL,经4层纱布过滤后直接测定pH并记录,然后将瘤胃液分装于冻存管中,于-20℃保存待测。瘤胃液中氨态氮(NH3-N)含量用苯酚-次氯酸钠比色法测定;瘤胃挥发性脂肪酸含量采用气相色谱法,使用气相色谱仪(GC-2014,岛津株式会社,日本)进行测定;瘤胃微生物蛋白(MCP)含量使用试剂盒进行测定,方法及步骤详见说明书,试剂盒购自南京建成生物工程研究所。
瘤胃液样品送上海美吉生物医药科技有限公司测定瘤胃菌群。根据E.Z.N.A.soil DNA kit(O-megaBio-Tek,美国)说明书进行微生物群落总基因组DNA抽提,完成样品DNA的抽提后,使用携带Barcode序列的上游引物338F(5′-ACTC-CTACGGGAGGCAGCAG-3′)和下游引物806R(5′-GGACTACHVGGGTWTCTAAT-3′)对16S rRNA基因V3~V4可变区进行PCR扩增,后将PCR扩增产物切胶回收,用Qubit4.0(Thermo Fisher Scientific,美国)对回收产物进行检测定量。使用NEXTFLEX Rapid DNA-Seq Kit对纯化后的PCR产物进行建库,最后利用Illumina NextSeq 2000平台进行测序。使用fastp软件对双端原始测序序列进行质控,使用FLASH软件进行拼接,然后使用Qi-ime2流程中的DADA2插件对质控拼接之后的优化序列进行降噪处理。根据ASV代表序列及丰度信息,进行群落多样性分析、菌群差异分析及相关性分析等。
委托上海美吉生物医药科技有限公司完成瘤胃液代谢组分析。本次液相色谱-质谱联用(LC-MS)分析的仪器平台为赛默飞公司的超高效液相色谱串联傅里叶变换质谱UHPLC-Q Exactive系统。将2μL样本经HSST3色谱柱(100 mm×2.1mm,1.8μm)分离后进入质谱检测。上机完成之后,LC-MS原始数据导入代谢组学处理软件Progenesis QI进行基线过滤、峰识别、积分、保留时间校正、峰对齐,最终得到一个保留时间、质荷比和峰强度的数据矩阵,同时将MS和MSMS质谱信息与代谢公共数据库HMDB和Metlin以及美吉自建库进行匹配,得到代谢物信息。
试验数据先用Excel 2019软件进行初步整理,后用SPSS 27.0进行独立样本t检验,结果数据以平均值和均值标准误的形式表示,P<0.05为差异显著,P<0.01为差异极显著,将0.05≤P<0.10作为有显著趋势。用 Wilcoxon 检验分析CON组和YC组的α多样性指数和2组在门水平和属水平上相对丰度的差异,将P<0.05定为差异显著性判断标准;用斯皮尔曼相关性分析对2组差异菌属和瘤胃发酵参数进行相关性分析。将美吉云平台上预处理后的矩阵文件进行差异分析。R软件包进行主成分分析。差异代谢物的选择基于正交偏最小二乘判别分析(OPLS-DA)模型,以重要变量投影(VIP)值>1、P<0.05的代谢物为差异代谢物。差异代谢物通过KEGG数据库进行的代谢通路注释,获得差异代谢物参与的通路。Py-thon 软件包scipy.stats进行通路富集分析。
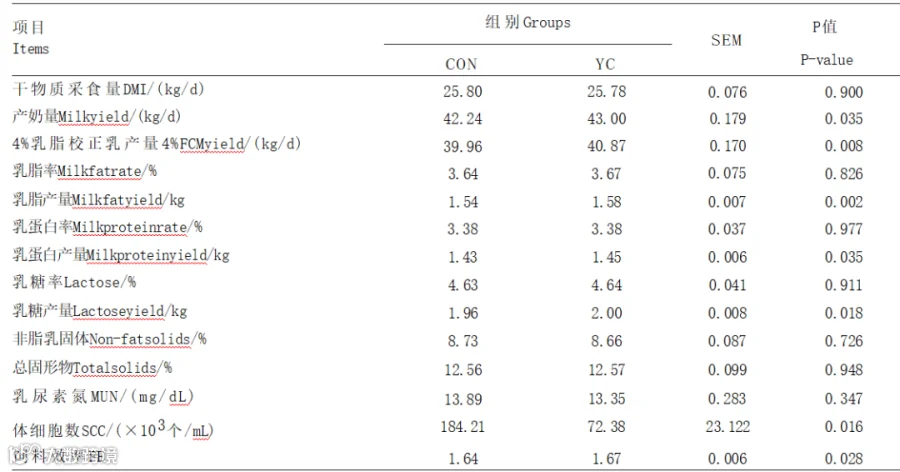
由表2可知,YC组的产奶量及4%FCM、乳脂、乳糖、乳蛋白产量显著高于CON组(P<0.05),YC组的体细胞数相较于CON组显著降低(P<0.05)。与CON组相比,YC组FE显著提高(P<0.05)。2组间奶牛的DMI、乳蛋白率、乳脂率、乳糖率及非乳脂固体、总固形物及乳尿素氮含量均无显著差异(P>0.05)。
表2YC对奶牛生产性能的影响
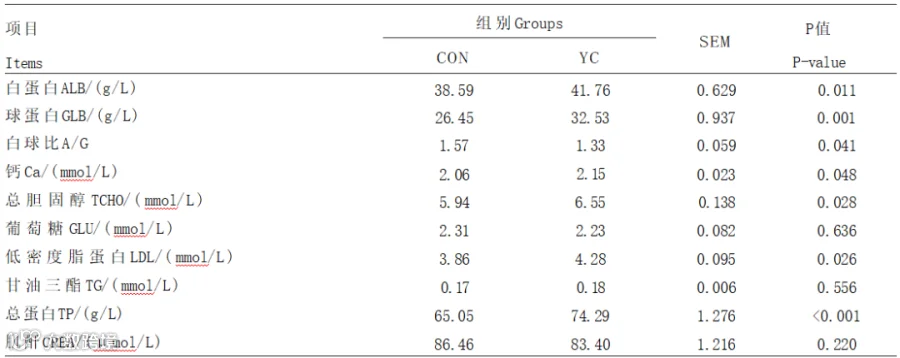
由表3可知,与CON组相比,YC组血清总蛋白含量极显著升高(P<0.01),血清白蛋白、钙、总胆固醇、低密度脂蛋白含量显著升高(P<0.05)。2组间奶牛的血清葡萄糖、甘油三酯和肌酐含量均无显著差异(P>0.05)。
表3 YC对奶牛血清生化指标的影响
由表4可知,YC组血清CAT和SOD活性极显著提高于CON组(P<0.01)。2组间血清GHS-Px、GST活性及MDA含量均无显著差异(P>0.05)。
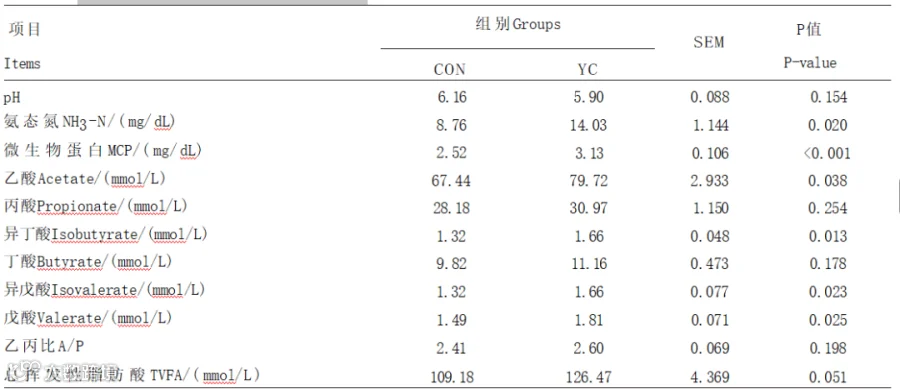
由表5可知,与CON组相比,YC组瘤胃液MCP含量极显著增加(P<0.01);YC组瘤胃液氨态氮、乙酸、异丁酸、异戊酸、戊酸含量显著提高(P<0.05),总挥发性脂肪酸含量有升高的趋势(P=0.051)。2组间奶牛瘤胃液丙酸、丁酸含量及乙丙比均无显著差异(P>0.05)。
表4 YC对奶牛血清抗氧化指标的影响
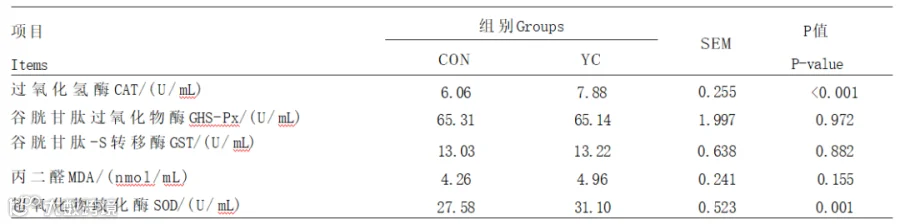
表5 YC对奶牛瘤胃发酵参数的影响
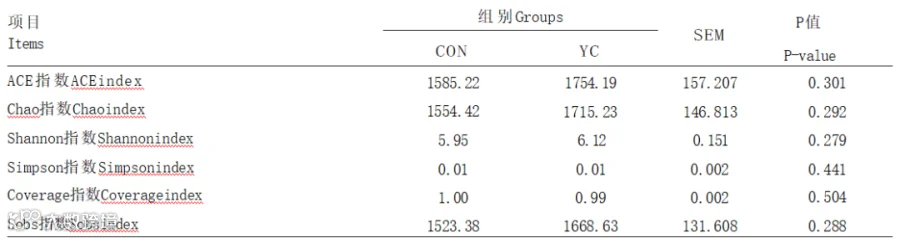
由表6可知,与对照组相比,添加YC对瘤胃液中微生物ACE、Chao、Shannon、Simpson、Cover-age、Sobs指数无显著影响(P>0.05),说明2组物种多样性无显著差异。
表6 YC对奶牛瘤胃微生物α多样性的影响
由图1-A可知,YC组和CON组的优势菌门均为拟杆菌门(Bacteroidota)、厚壁菌门(Firmi-cutes)、变形菌门(Proteobacteria),其中拟杆菌门的相对丰度最高。由图1-B可知,2组的优势菌属均为普雷沃氏菌属和解琥珀酸菌属。
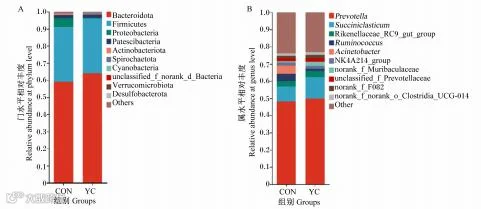
Bacteroidota:拟杆菌门;Firmicutes:厚壁菌门;Proteobacteria:变形菌门;Patescibacteria:髌骨菌门;Actinobacteriota:放线菌门;Spirochaetota:螺旋体菌门;Cyanobacteria:蓝藻菌门;Verrucomicrobiota:疣微菌门;Desulfobacterota:脱硫杆菌门;Prevo-tella:普雷沃氏菌属;Succiniclasticum:解琥珀酸菌属;Rikenellaceae_RC9_gut_group:理研菌科RC9_肠道群;Ruminococcus:瘤胃球菌属;Acinetobacter:不动杆菌属;NK4A214_group:NK4A214群;norank_f_Muribaculaceae:未分类拟杆菌目;unclassified_f_Prevotellaceae:未分类普雷沃氏菌科;norank_f_norank_o_Clostridia_UCG-014:未分类梭菌纲UCG-014;Other:其他。
图1 瘤胃菌群在门(A)和属水平(B)的群落分布
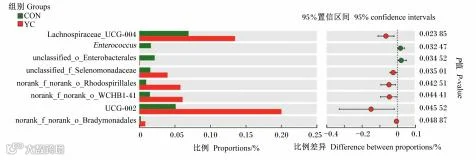
如图2所示,YC组中的6个属:UCG-002、毛螺菌科UCG-004、norank_f_norank_o_WCHB1_41、未分类红螺菌目、未分类硒单胞菌科、未分类慢生单胞菌目的相对丰度显著高于CON组(P<0.05);YC组中的未分类肠杆菌目、肠球菌属的相对丰度显著低于CON组(P<0.05)。
Lachnospiraceae_UCG-004:毛螺菌科UCG-004;Enterococcus:肠球菌属;unclassified_o_Enterobacterales:未分类肠杆菌目;unclassified_f_Selenomonadaceae:未分类硒单胞菌科;norank_f_norank_o_Rhodospirillales:未分类红螺菌目;norank_f_norank_o_Bradymonadales:未分类慢生单胞菌目。
图2 CON组和YC组差异菌属
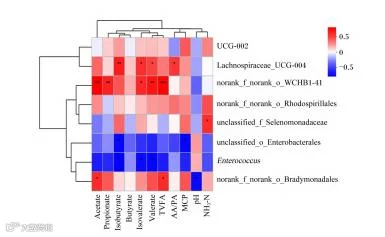
如图3所示,毛螺菌科UCG-004相对丰度与异丁酸、异戊酸、戊酸含量呈显著或极显著正相关(P<0.05或P<0.01)。norank_f_norank_o_WCHB1_41相对丰度与乙酸、丙酸、异戊酸、戊酸及总挥发性脂肪酸含量呈显著或极显著正相关(P<0.05或P<0.01)。未分类硒单胞菌科相对丰度与NH3-N含量呈显著正相关(P<0.05)。
Lachnospiraceae_UCG-004:毛螺菌科UCG-004;norank_f_norank_o_Rhodospirillales:未分类红螺菌目;unclassified_f_Sel-enomonadaceae:未分类硒单胞菌科;unclassified__o_Enterobacterales:未分类肠杆菌目;Enterococcus:肠球菌属;norank__f_norank_o_Bradymonadales:未分类慢生单胞菌目;Acetate:乙酸;Propionate:丙酸;Isobutyrate:异丁酸;Butyrate:丁酸;Isovaler-ate:异戊酸;Valerate:戊酸;TVFA:总挥发性脂肪酸totalvolatilefattyacids;AA/PA:乙丙比acetate/valerate;MCP:微生物蛋白microprotein;NH3-N:氨态氮ammoniacalnitrogen。
∗表示显著相关(P<0.05),∗∗表示极显著相关(P<0.01),∗∗∗表示极显著相关(P<0.001)。图7同。∗indicates sig-nificant correlation(P<0.05),∗∗indicates extremely significant correlation (P<0.01),and∗∗∗indicates extremely significant correlation (P<0.001).The same as Fig.7.
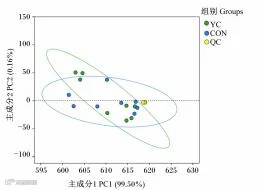
QC:质量控制样本qualitycontrolsample
由图4可知,PCA模型拟合程度较好,但添加YC对瘤胃液代谢物整体没有显著影响。
由图5可知,2组样品共鉴定出224个差异代谢物,上调155个,下调69个。与CON组相比,YC组共筛选到224种差异代谢物,其中三甲基吲哚、羟基苯乙酰甘氨酸、1-丁醇、N-羟基色胺、4-吡啶氧酸、2-甲基-3-羟基-5-甲酰吡啶-4-羧酸盐等代谢物含量显著上调(表7)。
2.6.3 差异代谢物的通路分析
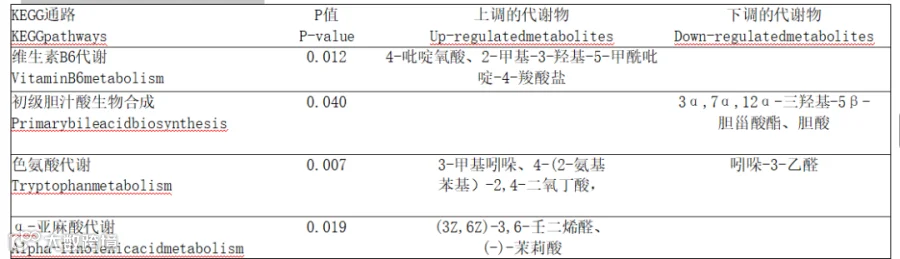
通过超几何检验分析,将P<0.05的代谢物进行KEGG代谢通路富集分析。由表8和图6可知,与CON组相比,YC组差异代谢物主要显著富集的通路为维生素B6代谢(vitamin B6 metabo-lism)、初级胆汁酸生物合成(primary bile acid bio-synthesis)、色氨酸代谢(tryptophan metabolism)以及α-亚麻酸代谢(alpha-linolenic acid metabo-lism)。
差异菌属与试验组差异代谢物的相关分析如图7所示。UCG-002相对丰度与N-羟基色胺、(3Z,6Z)-3,6-壬二烯醛含量呈显著正相关(P<0.05);norank_f_norank_o_WCHB1-41相对丰度与(S)-2,3,4,5-四氢二吡啶甲酸盐、羟基苯乙酰甘氨酸、1-丁醇、2-甲基-3-羟基-5-甲酰吡啶-4-羧酸盐含量呈显著或极显著正相关(P<0.05或P<0.01)。毛螺菌科UCG-004相对丰度与(S)-2,3,4,5-四氢二吡啶甲酸盐、3-甲基吲哚、4-吡啶氧酸、羟基苯乙酰甘氨酸、N-羟基色胺、1-丁醇、2-甲基-3-羟基-5-甲酰吡啶-4-羧酸盐含量呈显著或极显著正相关(P<0.05或P<0.01)。
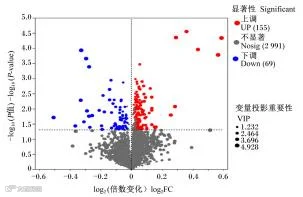
横坐标代表组间差异的倍数,纵坐标是代谢物表达量变化差异的统计学检验值,值越大差异越显著。每个点代表1个代谢物,点的大小代表重要变量投影值。红色点是显著上调的代谢物,蓝色点是显著下调的代谢物,灰色点则代表差异不显著的代谢物。

奶牛的产奶量和乳成分直接影响了牧场的经济效益。Dais等研究表明,添加YC可以显著提高产奶量、3.5%FCM产量和能量校正乳产量,还提高了乳脂和乳蛋白产量。研究表明,围产期添加活酵母可以显著提高产奶量,提高乳蛋白含量,减少体细胞数。添加YC和酶解酵母可以显著提高泌乳早期奶牛的产奶量、4%FCM产量和能量校正乳产量。Halfen等报道,泌乳中期添加YC对产奶量并无显著影响。
本试验中,添加YC显著提高了产奶量及4%FCM、乳脂、乳蛋白、乳糖产量及FE。YC提高产奶量,提升乳品质的原因一方面可能与其本身含有的可发酵代谢物有关,这部分代谢物直接被瘤胃细菌利用;另一方面YC通过影响瘤胃菌群结构来影响碳水化合物等营养物质在瘤胃中的发酵,进而提升能量利用效率,产生更多的牛奶。此外,本研究中发现添加YC还能显著减少体细胞数,这很可能由于YC中含有的甘露寡糖(MOS)和β-葡聚糖。MOS对有益菌具有促进作用,而对有害菌的定植和生长具有抑制作用,进而影响瘤胃营养物质的降解,从而提高了动物的生产性能和抗病能力,同时,MOS还可以作为免疫刺激的辅因子,直接作用于机体的免疫系统,提高动物的免疫力。而β-葡聚糖在很多报道中已经证明了其对动物在抗氧化和免疫方面的有效性。
血清生化指标在一定程度上能够反映机体的生理状态。血液中白蛋白可以维持血液的渗透压,运输多种代谢物。球蛋白由免疫器官生成,与机体免疫力密切相关。总蛋白是白蛋白和球蛋白之和。本试验中,添加YC显著提高了血清中总蛋白的含量,这可能与YC提升了奶牛对蛋白质消化吸收与利用的能力有关。
胆固醇是细胞膜结构的重要组成部分,对于细胞膜稳定性和流动性维持具有重要作用。胆固醇还可以生成胆酸,参与脂肪消化。此外,胆固醇还是机体中类固醇激素合成的前提物质。低密度脂蛋白是血浆中胆固醇占比最高的一类脂蛋白,为蛋白质、胆固醇和磷脂组成的复合体,是极低密度脂蛋白降解的产物,主要作用是运载胆固醇进入外周组织细胞。李琛研究表明,在饲粮中添加YC可以显著提高奶牛血清中低密度脂蛋白和胆固醇的含量。本试验中,添加YC显著提高了血清低密度脂蛋白和胆固醇含量,在一定程度上说明机体对脂肪酸的利用能力得以提升,这与前人的试验结果是一致的。
表7 2组瘤胃液显著差异代谢物
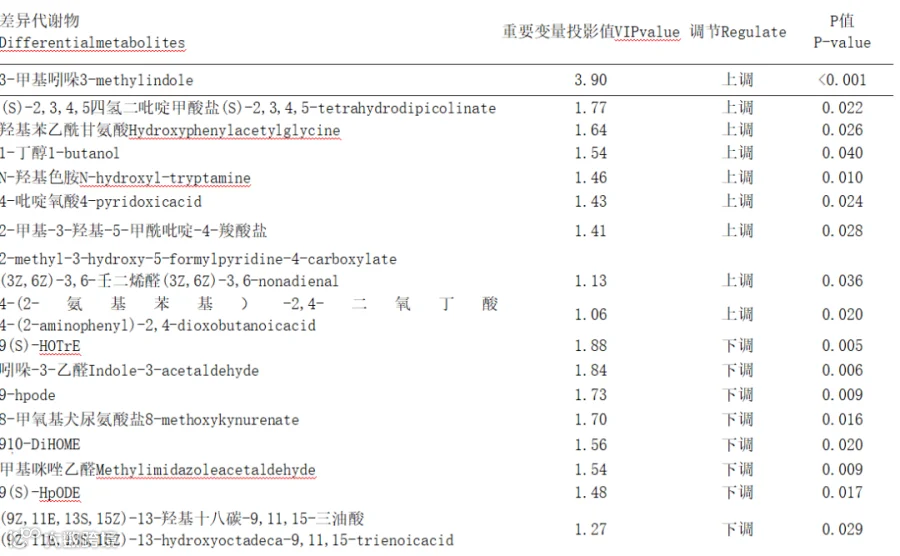
表8 2组瘤胃液KEGG代谢通路
有研究表明,由于自由基的过度生成导致机体处于氧化应激状态。CAT是一种重要的抗氧化酶,其主要作用是催化在氧化还原过程中产生的过氧化氢,以减少过氧化氢对细胞的氧化损伤。SOD是一种天然的抗氧化金属酶,具有清除活性氧、缓解炎症和抗氧化等生物学功能。王卫正探究了在饲粮中添加YC对泌乳初期奶牛抗氧化功能的影响,研究结果表明,在饲粮中添加3%YC极显著提高了血液中SOD的活性。本试验结果表明,添加YC可以显著提升血清CAT和SOD的活性,提升了奶牛的抗氧化能力,这与前人研究结果一致。程艳研究表明,添加YC可以显著提高隐形乳房炎奶牛血浆中GSH-Px活性,本试验中血清GSH-Px的活性在YC组并无显著升高,这可能与奶牛的健康状态有关。
瘤胃NH3-N由瘤胃菌群分解含氮物质的产生,同时微生物利用NH3-N合成MCP。充足的NH3-N是保证MCP合成的前提。郭婷研究表明,添加YC会通过影响瘤胃中纤维降解菌、蛋白水解菌以及淀粉降解菌等细菌之间的相互作用从而提高瘤胃中NH3-N和MCP的含量,这与本试验结果一致。

Vitamin B6 metabolism:维生素B6代谢;Primary bile acid biosynthesis:初级胆汁酸生物合成;Tryptophan metabo-lism:色氨酸代谢;Alpha-linolenicac idmetabolism:α-亚麻酸代谢。
图中每个气泡代表1个代谢通路;横轴表示通路中代谢物相对重要性;纵轴表示代谢物参与通路的富集显著性;
气泡大小代表通路的重要性;气泡越大,通路重要性越大。Each bubble in the figure represents a metabolic pathway;the transverse axis indicates the relative importance of metabolites in the pathway;the longitudinal axis indicates the enrichment significance of metabolites involved in the pathway;the bub-ble size represents the importance of the pathway;the larger the bubble,the greater the importance of the pathway.
图6 瘤胃液差异代谢物通路分析
乙酸是瘤胃发酵的终产物,是脂肪酸从头合成的主要底物;增加乙酸供应会提高产奶量和乳脂产量。有研究表明,在饲粮中添加活性干酵母可以改善瘤胃发酵,即提高瘤胃中乙酸和丁酸的含量。瘤胃中丙酸含量受饲粮中精料比的影响,一般情况下精料比越高产生的丙酸也越多。丙酸是肝脏糖异生的主要原料,是奶牛能量的重要来源。丁酸主要由存在于反刍动物消化道中的厚壁菌门分解碳水化合物生成,丁酸可以作为合成乳脂和体脂的前提物质,也可以参与机体供能,另外,丁酸还可以生成胆固醇。异位酸含有支链挥发性脂肪酸,瘤胃中可以产生的异位酸主要包括异戊酸、异丁酸和2-甲基丁酸,支链挥发性脂肪酸是瘤胃纤维素分解菌生长的必需营养素。在本研究中,与CON组相比,YC组中的乙酸、异戊酸、异丁酸、戊酸含量显著升高,这与Lascano等研究的结果是一致的。本试验中,乙酸含量的升高可能因为YC刺激了纤维素分解菌的生长。张少泽研究表明,在奶牛饲粮中添加YC对瘤胃中乙酸含量无显著影响,与本试验结果不一致,可能与奶牛的饲粮组成、泌乳阶段、生理状态以及所处的环境等因素相关。
在本研究中,2组的优势菌门都为拟杆菌门、厚壁菌门、变形菌门,其中拟杆菌门的相对丰度最高。Xue等通过微生物基因组说明了拟杆菌门在瘤胃碳水化合物降解过程中发挥了重要作用。Xue等研究表明,厚壁菌门具有促进纤维在瘤胃中降解的功能。2组的优势菌属均为普雷沃氏菌属。这与前人的研究结果一致。普雷沃氏菌属对反刍动物瘤胃降解碳水化合物具有重要作用,其在瘤胃中的相对丰度可间接说明瘤胃对碳水化合物的消化水平,此外,普雷沃氏菌属也对蛋白质的降解发挥重要作用。本试验中,YC组中的UCG-002、毛螺菌科UCG-004、未分类硒单胞菌科等的相对丰度显著高于CON组,这些菌属都属于厚壁菌门。所以,添加YC可能促进纤维在瘤胃中的降解,这也可能是YC组中挥发性脂肪酸含量升高的原因。试验组中的norank_f_norank_o_WCHB1_41属于疣微菌门,一些疣微菌对多糖和木质纤维素的降解具有重要作用。在本试验中,norank_f_norank_o_WCHB1_41相对丰度与乙酸、丙酸、异戊酸等挥发性脂肪酸及总挥发性脂肪酸含量呈显著或极显著正相关。UCG-002属于颤螺菌科,Gophna等在研究中表明,颤螺菌属可以发酵碳水化合物。本试验的相关性结果表明,毛螺菌科UCG-004相对丰度与异丁酸、异位酸等异位酸含量的升高有显著或极显著的正相关关系,毛螺菌科UCG-004属于毛螺菌科,有研究表明,毛螺菌属可以发酵各种复杂的多糖,产生挥发性脂肪酸,而且毛螺菌科还可以利用氨基酸合成异位酸。这和本研究中试验组乙酸、异丁酸、异戊酸含量的显著升高结果一致。试验组中的未分类硒单胞菌科属于硒单胞菌科,硒单胞菌可以对多种碳水化合物分解代谢,还可以利用蛋白质、氨和氨基酸作为氮源,并利用氨进行氨基酸的生物合成。与CON组相比,添加YC还显著降低了试验组中未分类肠杆菌目和肠球菌属等可能致病的菌属的相对丰度,这可能与酵母细胞壁可以抑制有害菌的繁殖有关。
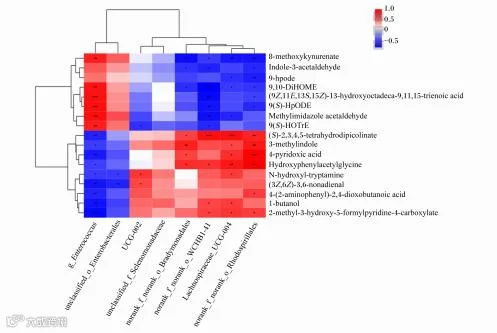
图7 差异代谢物与差异菌属相关性分析
g_Enterococcus:肠球菌属;unclassified_o_Enterobacterales:未分类肠杆菌目;unclassified_f_Selenomonadaceae:未分类硒单胞菌科;norank_f_norank_o_Bradymonadales:未分类慢生单胞菌目;Lachnospiraceae_UCG-004:毛螺菌科UCG-004;norank_f_norank_o_Rhodospirillales:未分类红螺菌目;8-methoxykynurenate:8-甲氧基犬尿氨酸盐;Indole-3-acetaldehyde:吲哚-3-乙醛;(9Z,11E,13S,15Z)-13-hydroxyoctadeca-9,11,15-trienoic acid:(9Z,11E,13S,15Z)-13-羟基十八碳-9,11,15-三油酸;Methylimidazole acetaldehyde:甲基咪唑乙醛;(S)-2,3,4,5-tetrahydrodipicolinate:(S)-2,3,4,5-四氢二吡啶甲酸盐;3-methylindole:3-甲基吲哚;4-pyridoxic acid:4-吡啶氧酸;Hydroxyphenylacetylglycine:羟基苯乙酰甘氨酸;N-hydroxyl-tryptamine:N-羟基色胺;(3Z,6Z)-3,6-nonadienal:(3Z,6Z)-3,6-壬二烯醛;4-(2-aminophenyl)-2,4-dioxobutanoicacid:4-(2-氨基苯基)-2,4-二氧丁酸;1-butanol:1-丁醇;2-methyl-3-hydroxy-5-formylpyridine-4-carboxylate:2-甲基-3-羟基-5-甲酰吡啶-4-羧酸盐。
瘤胃代谢组学分析结果表明,显著差异的通路分别为维生素B6代谢、初级胆汁酸生物合成、色氨酸代谢、α-亚麻酸代谢。维生素B6是一种水溶性维生素,参与机体蛋白质代谢、糖原与脂肪酸代谢,参与一些微量营养素的转化与吸收,调节神经递质的合成和代谢。黎俊报道,维生素B6是蛋白质代谢过程中氨基酸转氨酶和脱羧酶的重要辅酶。脱羧作用能够促进神经递质的合成,如多巴胺、组胺等。通过转氨酶调控色氨酸代谢,色氨酸也可以通过相关途径合成烟酸。色氨酸不但在蛋白质生物合成具有重要作用,而且它还是神经递质的前体,如烟酰胺、维生素B6等。此外,色氨酸还可以提高动物的抗氧化和免疫力。α-亚麻酸是一种多不饱和脂肪酸,具有抗炎、抗氧化作用。上述代谢通路说明添加YC会影响瘤胃维生素代谢、氨基酸代谢和脂代谢。

饲粮中添加YC可以提高奶牛瘤胃特定细菌丰度,增加挥发性脂肪酸、MCP和B族维生素等合成,改善瘤胃发酵效率并增强奶牛抗氧化能力,进而提高泌乳后期奶牛的生产性能。
文章来源自:动物营养学报 2025,37(7):4576⁃4590Chinese Journal of Animal Nutrition,作者,米虎伟 李一帆 王 丽 李玉龙 曹雨娃 王弘浩 刘维平王腊梅 姚军虎 王砀砀 曹阳春

鹤来生物科技(济南)
电话:18963489690