

一说到肠道菌群,大家第一感觉可能就是在我们肠道里上百种、上万亿微生物组成的复杂生态系统。不论是在动物界还是在人医的学术界乃至在大众的认知中,对“菌群平衡”有个根深蒂固的认知。
第一:菌群的种类越多,分布越均匀,肠道就越健康,学术上我们叫做α多样性,通俗理解就是肠道里细菌的种类够不够丰富,各类菌的数量分布是不是相对均匀,通常菌群越多越均匀我们就自然而然的认为这样的肠道更健康。
第二:菌群出问题,根源在于“好菌少了,坏菌多了”。按照这个逻辑,我们只要找到疾病中哪些菌异常升高,哪些菌异常降低,是不是就找到了疾病的生物标志物,甚至可以直接作为治疗的靶点。
这两个判断听起来合理,也指导了过去二十年的菌群研究。但是当一组研究人员系统性地回顾了炎症性肠病、艰难梭菌感染、肠易激综合征、结直肠癌这四大类肠道疾病的临床宏基因组数据,结果揭示了一个令人不安的事实:炎症性肠病和艰难梭菌感染患者的菌群多样性确实明显低于健康人群,但肠易激综合征和结直肠癌患者的菌群多样性,跟健康人相比竟然没有统计学上的显著差别。
这意味着什么?多样性下降根本不能作为判断肠道疾病的通用标准。 有些人的肠道菌群看上去种类齐全、分布匀称,但实际上菌群网络已经乱了套,甚至在毫无察觉的情况下出现了早期癌变。
更让人头疼的是,不同肠道疾病中异常变化的菌种彼此之间几乎没有重叠。你想找一个能同时区分健康人和多种肠道疾病患者的“通用菌种标志物”?根本找不到。这正是为什么直到今天,“菌群失调”这四个字仍然没有一个统一、清晰的定义;这也解释了为什么益生菌干预、粪菌移植这些疗法,在不同的患者身上效果天差地别,几乎无法提前预测谁会受益、谁不会有反应。
问题究竟卡在哪里?研究团队给出的回答直击要害:过去所有人的目光都聚焦在“肠道里住着哪些菌”,却完全忽视了“这些菌彼此之间在发生什么样的关联”。而真正决定一个微生物生态系统是健康还是病态的,恰恰是后者。

为了搞清楚肠道菌群动态变化的底层逻辑,研究团队构建了一个代谢明确模型,完整模拟了肠道里细菌的生长、代谢、繁殖,以及它们之间的相互作用。
这个模型最大的价值,是它没有提前预设任何结果,而是完全基于细菌的代谢规律,让菌群的群落结构自然形成。最终,模型自发演化出了两种完全不同、且极其稳定的生态状态,而这两种状态,恰好完美匹配了真实世界里健康肠道和失调肠道的所有特征。
这两种状态的核心区别,通俗理解起来:

在健康的肠道生态里,细菌之间最核心的关系,是互相竞争。它们会争抢肠道里的营养物质、生存空间,互相制约,谁也无法一家独大。
这种竞争带来了两个核心的健康价值:
维持了系统的韧性:没有任何一种菌能疯狂繁殖,整个菌群的结构始终保持动态稳定,就算有外界干扰(比如偶尔吃一顿重油重盐的食物、吃了抗生素),也能快速恢复平衡。
保留了充足的功能冗余:不同的细菌会进化出重叠的代谢功能,就像一个公司里,同一个岗位有多个备选人员,就算某一种菌的数量下降了,其他菌也能立刻补上它的功能,不会让整个肠道的代谢链条断裂。

在失调的肠道生态里,情况完全反转了:细菌之间的核心关系,变成了互利共生、交叉摄食。这里的交叉摄食,通俗说就是细菌之间形成了一条闭环的代谢食物链:A菌分解肠道里的营养物质,排出的代谢废物刚好是B菌的口粮;B菌代谢产生的物质,又刚好能喂饱C菌;而C菌的产物,又能反过来被A菌利用。
这群抱团合作的细菌,形成了一个自给自足、效率极高的菌群联盟,它们能快速抢占肠道里的所有资源,疯狂繁殖,最终彻底主导整个肠道生态。而这种状态,带来了灾难性的后果:
菌群联盟会挤走其他无法融入闭环的细菌,最终导致肠道的功能冗余丧失,整个系统的抗干扰能力大幅下降;
这个高效的代谢闭环,会过度消耗肠道里的营养,甚至产生对宿主有害的代谢产物,直接伤害肠道黏膜,引发慢性炎症,甚至推动癌变;
这种抱团的稳态极其顽固,一旦形成就很难打破,这也是为什么很多慢性肠道疾病反复发作、难以根治。

瘤胃本质上就是一个厌氧发酵罐,其中栖息着细菌、原虫、厌氧真菌和产甲烷古菌构成的复杂微生物群落。这个微型生态系统与肠道菌群共享着完全相同的生态学底层逻辑——它们都是微生物相互作用网络。
当我们把Science研究的生态学框架移植到瘤胃上,很多长期困扰反刍动物营养实践的问题,突然有了全新的解读视角。
以瘤胃酸中毒为例,过去我们一直把目光盯在pH值上。但从微生态视角看,这不仅是化学指标的波动,更是一场菌群相互作用网络的失衡。

在健康的瘤胃里,乳酸产生菌(以牛链球菌为代表)和乳酸利用菌(以埃氏巨型球菌、反刍兽新月单胞菌为代表)之间,维持着一种经典的竞争与制约关系。
这两种菌并不直接争抢同一种食物——乳酸产生菌吃的是碳水化合物,乳酸利用菌吃的是乳酸。但它们之间的竞争是真实存在的,体现在两个层面:其一,对瘤胃内有限生态空间的争夺;其二,乳酸产生菌的代谢活动会降低pH,而乳酸利用菌对低pH更敏感——这意味着,乳酸产生菌的增殖本身,就是在恶化乳酸利用菌的生存环境。
这种竞争之所以被称为“滞后竞争”,是因为乳酸利用菌的增殖天然慢于乳酸产生菌。乳酸产生菌繁殖快、产酸快,占据先发优势;乳酸利用菌繁殖慢,且必须以乳酸为底物,只能在乳酸产生菌“先行动”之后才能响应。在健康的瘤胃里,由于碳水化合物不会过量涌入,乳酸产生菌的扩张是温和的,乳酸利用菌虽有滞后,但仍有足够的时间追上节奏、将乳酸持续清除。两者之间形成了一种动态制衡——乳酸产生菌无法无限扩张,乳酸利用菌也不会被完全压制。这让我想到了为什么养殖户喂牛时需要控制采食时间的一个原因,饲草型日粮的牛可以一直吃到他们的瘤胃填满为止。相反,以谷物为基础的日粮尤其是高精料饲喂模式下的牛每次只能摄入有限的量,从而在下一进食之前有时间分解积累的酸,这也就是为什么控制采食时间的问题。

当高精料日粮涌入大量易发酵碳水化合物时,乳酸产生菌在短时间内暴增,乳酸产量远超乳酸利用菌的处理能力。pH迅速下降,乳酸利用菌自身的活性被抑制。
在这个节点上,一种更隐蔽的生态学转变发生了:当瘤胃pH持续走低,一部分耐酸菌群(包括特定乳酸杆菌菌株、耐酸的埃氏巨型球菌等)开始占据生态位。它们与持续产酸的菌群之间,不再维持竞争制约关系,而是形成了交叉摄食的代谢闭环。
这个闭环的运作方式与Science研究所揭示的失调状态完全一致:产酸菌分解碳水化合物产生乳酸和部分代谢中间产物;耐酸菌恰好以这些中间产物和乳酸为底物,进一步代谢产生其他有机酸;而耐酸菌的代谢产物,又能够被产酸菌再次利用。这是一个自给自足、效率极高的“菌群联盟”。它不再依赖宿主日粮中原始的纤维或淀粉结构,而是依靠成员之间相互“喂养”来维持运转。这个闭环一旦形成,就会持续产酸、持续消耗营养、持续排挤其他无法融入闭环的菌群。

这正是Science研究所描述的失调状态的瘤胃版本。原文的消费者-资源模型揭示:在失调状态下,细菌群落通过交叉摄食形成高效的闭环,导致功能冗余丧失、系统韧性崩溃,而瘤胃酸中毒的本质,恰恰就是产酸菌与耐酸菌之间形成了交叉摄食闭环,替代了原本乳酸产生菌与乳酸利用菌之间“滞后竞争”的动态制衡关系。
表面上看,酸中毒是“乳酸堆积过多”;但从生态网络的视角看,它是菌群之间的相互作用模式的改变,从以竞争为主导的健康网络,滑向了以交叉摄食合作为主导的病态闭环。
这也解释了为什么单纯“补菌”难以从根本上解决酸中毒问题,只要这个交叉摄食闭环没有被打破,新补充进来的菌群就只能被排挤出局,解决问题的关键,不在于增加某一类菌的数量,而在于拆解这个闭环,重建竞争制衡的生态网络。
而这也正是酵母培养物——尤其是深度厌氧发酵酵母培养物能够发挥独特价值的切入点。



理解了酸中毒的生态学本质,酵母培养物的作用机制就不单单是简单的“益生元机制”,而是一套针对性的瘤胃菌群生态网络的修复方案。
第一:选择性促进乳酸利用菌,强化竞争地位。
酵母培养物对乳酸利用菌具有显著的选择性促生作用。已有研究表明,酵母培养物能够显著促进反刍兽新月单胞菌和埃氏巨型球菌的增殖。当这些分解乳酸的关键菌群获得营养支持、增殖速度加快时,它们在瘤胃生态网络中的竞争地位就得到了强化——它们能够更快地响应乳酸浓度的上升,及时将乳酸转化为挥发性脂肪酸,使乳酸的产生与清除重新回到动态平衡的轨道上。
第二:通过代谢产物调控,降低乳酸产生菌的产酸活性
深度厌氧发酵产生的酵母培养物富含多种微生物代谢产物。已有研究表明,酵母培养物的添加能够显著降低瘤胃中异常代谢产物的浓度,其中包括乳酸、脂多糖和组胺。这一抑制作用可能是存在多种层面上的机制:
首先,酵母培养物在发酵过程中产生的有机酸本身就具有抑制有害菌生长的作用。其次,酵母培养物在特定代谢条件下能够分泌具有抗菌活性的多肽类物质。此外,我们推测深度厌氧发酵技术的酵母培养物中可能含有大量用于菌群间沟通、竞争和防御的次级代谢物质,某些活性组分还可能通过干扰菌群间的群体感应信号,间接影响乳酸产生菌的代谢行为,但是具体的作用通路仍有待进一步研究阐明。

基于以上分析,我们可以给出一个更精确的表述:
酵母培养物对菌群平衡的作用,不是在“有益菌”和“有害菌”之间做简单的加减法,而是介入菌群之间的相互作用网络,重建乳酸产生菌和乳酸利用菌之间的竞争制衡关系,打破产酸菌与耐酸菌之间的交叉摄食闭环,使瘤胃生态系统回归到以竞争为主导、具备充足韧性与功能冗余的健康状态。
在这个框架下,益生元的价值——提供甘露聚糖、β-葡聚糖、小分子肽、核苷酸等营养底物,仍然是基础。它为乳酸利用菌的选择性促进提供了物质前提。但真正让深度厌氧发酵酵母培养物超越一般益生元的,是它能够进一步介入菌群之间的互作网络,推动整个生态网络的结构稳态。
这,就是鹤来理解的“菌群稳态平衡”——不是“一团和气”的静态和谐,而是充满良性竞争与相互制约的动态的稳态平衡。


并非所有酵母培养物都具备上述的菌群调控能力,浅层次发酵的酵母培养物的作用更多存在于初级营养代谢产物发挥的作用,但能否在益生元的基础上同时实现代谢信号的精准调控——即从“养菌”跨越到“调控菌群稳态”的更高维度,这取决于发酵工艺的深度。
要理解这一点,首先需要澄清一个关键概念:酵母培养物本质上是一种微生态制品,是在特定工艺条件下由酵母菌在特定培养基上经过充分厌氧发酵后形成的。它的价值从来不在于“活菌数”,而在于发酵过程中累积的代谢产物群。 大量研究证实,单纯依靠活酵母菌无法达到酵母培养物的效果。真正的差异化,藏在发酵工艺的深浅之间。
酵母是典型的兼性厌氧微生物,其代谢路径会随着氧气条件而根本性切换:在有氧条件下,酵母主要进行有氧呼吸,快速繁殖菌体;在厌氧条件下,则转向发酵途径,大量合成醇类、有机酸等代谢产物。这条代谢规律,决定了不同发酵深度下产物构成的本质差异:

浅层发酵或好氧发酵的核心特征是菌体繁殖主导。在有氧条件下,酵母优先利用氧气进行有氧呼吸,代谢通路主要服务于细胞增殖—大量合成菌体蛋白,将碳源和能量用于自身增殖。此时产生的代谢产物以初级代谢产物为主,种类有限、功能性单一。这种工艺虽然菌体产量高,但产物构成相对单薄,更偏向“蛋白饲料”而非“功能调控”。
从工艺实操层面看,市面上的浅层发酵产品普遍将发酵周期控制在7至10天左右。这背后有一个微生物生态学的现实制约:在发酵初期,酵母菌凭借其对氧气和糖分极强的利用效率,能够迅速占据优势,挤压其他微生物的生存空间。然而,由于浅层发酵并非严格厌氧环境,约48小时后,其他杂菌便开始抬头,逐步取代酵母的生态位。为避免杂菌污染和产品质量失控,生产方往往只能在发酵48-72小时后进行高温烘干,强制终止发酵进程。
这一工艺选择带来了两个连锁问题:
其一,内容物释放受限。 高温烘干导致酵母菌迅速死亡,但其细胞膜和细胞壁结构保持完整。这种物理屏障会阻碍动物消化道内的消化酶与酵母内容物(蛋白质、核酸、维生素等)直接接触,导致营养物质的生物利用率大打折扣。为弥补这一缺陷,生产上通常需要额外增加定向酶解或物理破壁工序来释放内容物,但这不仅增加了工艺复杂度,实际效果仍难以媲美酵母在自然衰亡裂解过程中释放产物的活性。
其二,名义添加成本与实际使用成本倒挂。 这类产品虽然名义单价较低,但由于功能性成分浓度有限、调控力不足,实际应用中的推荐添加量往往高达2%至4%,折算下来的单位使用成本并不经济,未能真正实现增效降本的初衷。

当发酵从浅层走向深层、从有氧切换为厌氧,酵母的代谢行为发生根本性转变:菌体增殖不再是唯一目标,取而代之的是次级代谢产物的合成。次级代谢产物是微生物在菌体生长进入稳定期后,以初级代谢产物为前体合成的功能性物质。这些产物与菌体生长繁殖无直接关系,但往往具备重要的生物活性—如抗生素、生物碱、生长因子和菌群信号分子。
正如发酵动力学所揭示的规律:次级代谢产物的合成受多种因素精密控制,需要在严格的厌氧环境和时间窗口下才能充分激活。深度厌氧发酵所提供的,正是这样一个完整而充分的“时间与空间”——让酵母走完从初级代谢到次级代谢的全过程,让活性信号物质得以充分积累。

我们对发酵第7天、第15天和第45天的代谢物进行了系统性检测与比较分析,发现发酵时间对代谢产物的种类、丰度和批次稳定性有着决定性的影响。从以下几个维度呈现核心研究结果。
a. 整体代谢轮廓:45天建立稳态,批次一致性最优
通过主成分分析(PCA)对三个时间点的整体代谢轮廓进行比较,结果如图1所示。D7样品点相对集中但仍有离散,反映快速增殖期的代谢波动;D15样品点离散度显著增大,表明发酵中期代谢网络尚未稳定,样本间个体差异明显;而D45样品点最为集中,4个重复样本的代谢物组成高度一致。
这一结果意味着,45天深度发酵不仅丰富了代谢产物的种类,更重要的是确保了批次间的可重复性—每一批产品都是可预期、可验证的“标准品”,而非充满随机波动的。
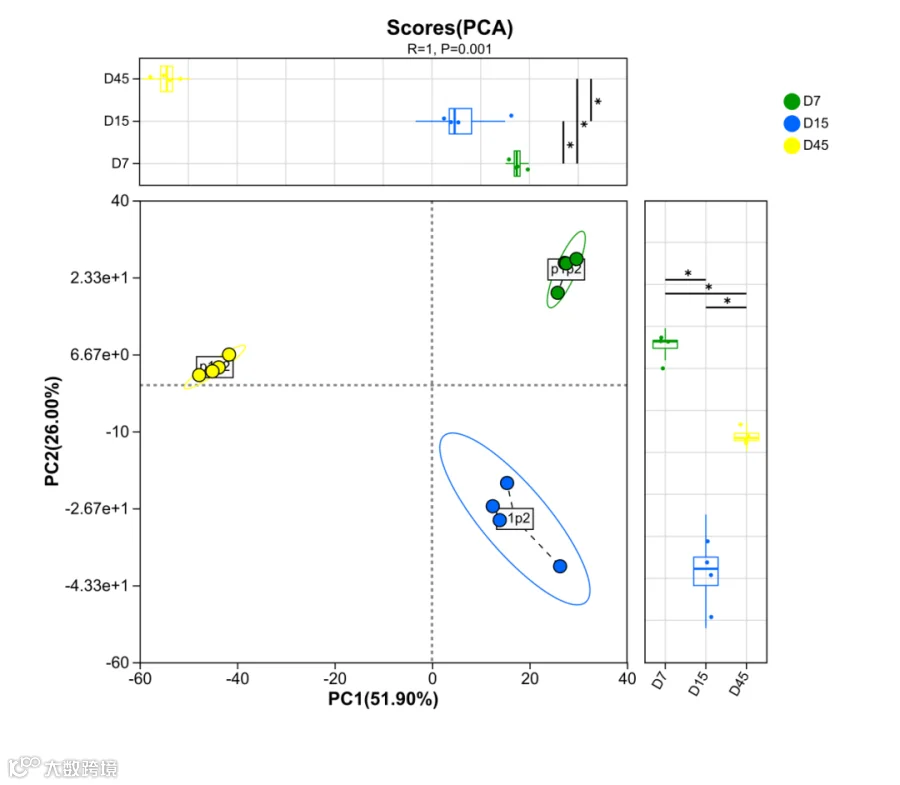
图1 D7、D15、D45整体比较
b. 有益代谢物的定向富集:4种关键物质持续上升
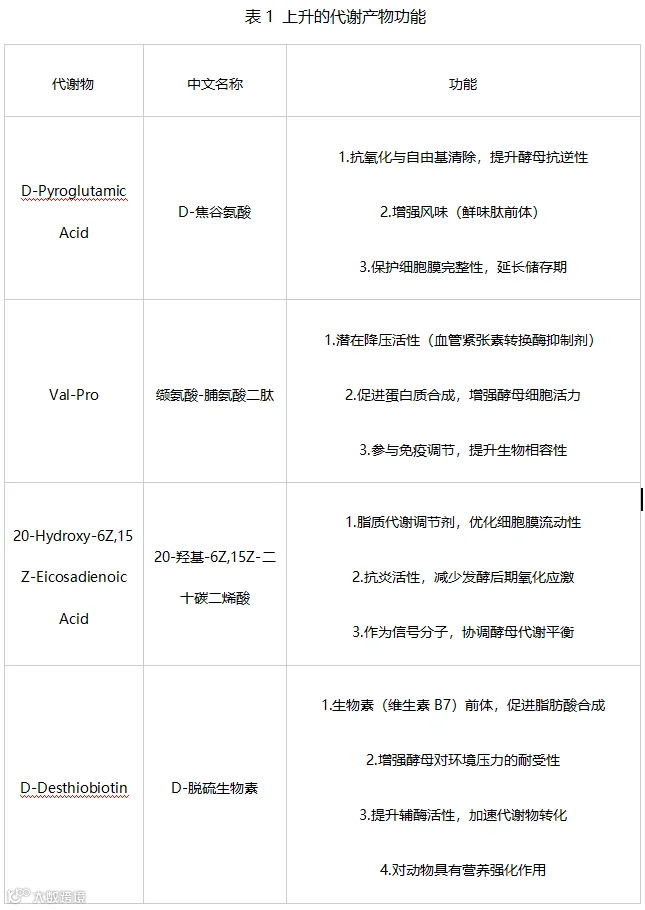
在45天发酵过程中,有4种代谢物呈现持续且显著的上升趋势(P<0.05),其变化轨迹如图2所示。这4种代谢物的具体功能见表1。
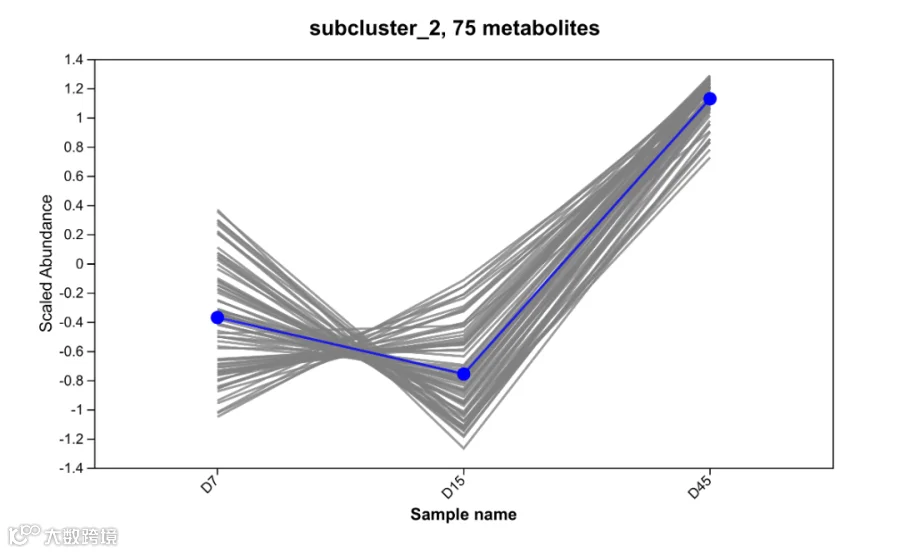
图2 持续上升代谢物趋势图
根据表1可知,这些代谢物在整个发酵过程显著上升,表明酵母进入成熟稳定期,代谢活动从快速增殖转向功能性物质合成,为酵母的营养与应用价值奠定基础。
c. 冗余及潜在有害物质的持续清除:20种代谢物显著下降
研究检测到20种代谢物在整个发酵过程中持续下降(P<0.05),其变化趋势如图3所示。表2选取其中具有代表性的几种代谢物,说明其下降的生物学意义。
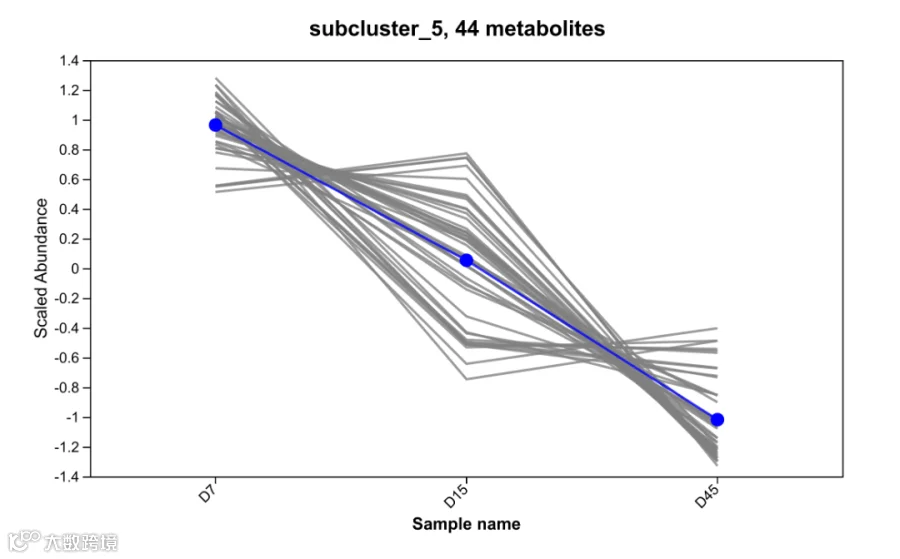
图3 持续下降代谢物趋势图

d. 综合以上数据,可以清晰地将酵母发酵过程划分为三个代谢阶段:
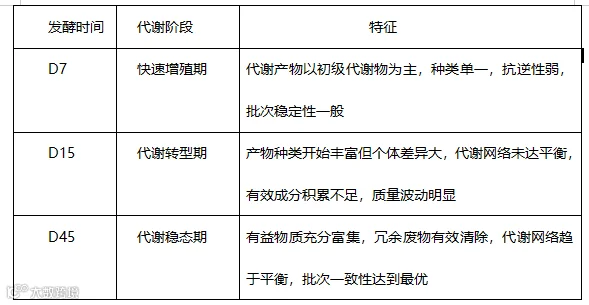
正如本研究的核心结论所言:酵母发酵45天优于7天/15天是代谢组的全面优化——有益物质富集、代谢废物清除、代谢效率提升、稳态成熟建立,产品质量稳定。这种代谢组学层面的成熟,正是45天发酵酵母在营养、功能、稳定性和经济性上全面超越短期发酵产品的根本原因。
深度厌氧发酵带来的不仅是代谢产物的种类和数量差异,更是产品稳定性的根本保障。鹤来生物采用的1080小时深度固态厌氧发酵工艺配合复合菌协同增效模式和高度受控的固体罐式发酵——充分释放了酵母从增殖、稳定到衰亡全生命周期的代谢潜力。优秀稳定的原料比例与安全可控的发酵环境,保障了酸度、酸溶蛋白占比和甘露寡糖等关键指标的稳定达标,使产品达到“湿料不霉变、不胀包”的品质状态,安全性与保质性行业领先。
这,就是鹤来生物二十余年来坚持走深度厌氧发酵之路的根本原因。我们选择了更难的路——更长的发酵周期、更复杂的工艺控制、更高的设备投入——因为只有深度厌氧发酵,才能让酵母完成从简单的“养菌”到更高层次的“菌群稳态平衡调控者”的角色跃迁。1080小时的发酵,换来的不仅是丰富的代谢产物组合,更是一个能够真正介入瘤胃菌群相互作用网络、推动系统从失衡回归稳态的“生态调控工具”。
结语:
酵母培养物的价值认知,应当从“益生元”的基础层面,提升到“菌群稳态平衡”的更高维度。鹤来生物的使命,就是通过不断精进深度厌氧发酵工艺,充分释放酵母培养物中的活性物质,帮助反刍动物的瘤胃建立起一个以竞争为主导、具备充足韧性和功能冗余的菌群稳态平衡。
上述引用文献为:
https://doi.org/10.1126/science.ady1729

部分内容参考微信公众号:生态学时空
