

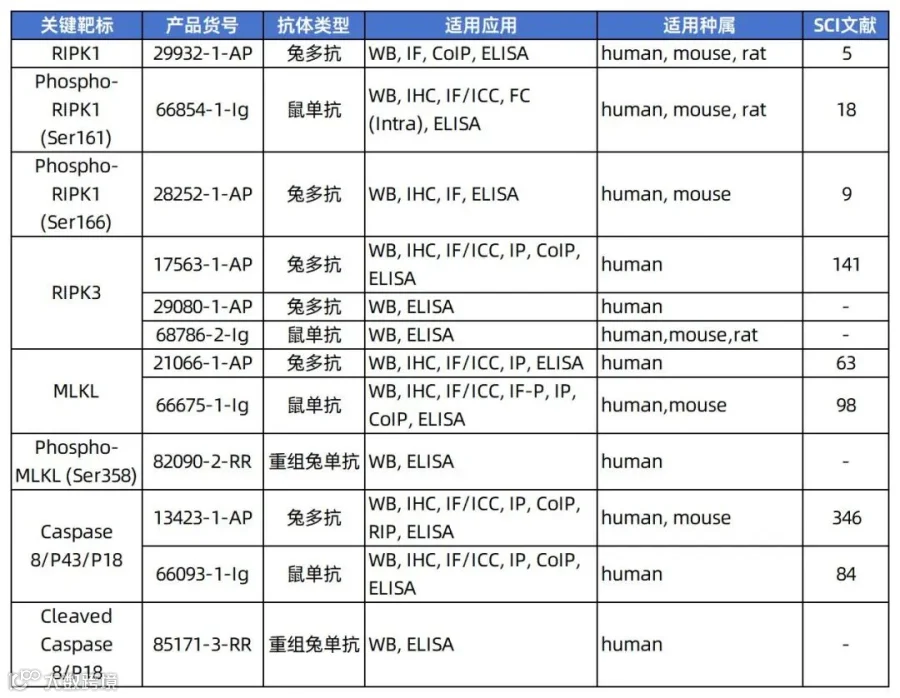
作为上期的延续(上期内容回顾),本期将介绍焦亡、铜死亡、双硫死亡、失巢凋亡、泛凋亡五种细胞死亡方式,及其关键靶标的实验技巧,篇幅较长,可跳转到感兴趣的主题哦~
1、什么是细胞焦亡?
(上下滑动查看全部)
细胞焦亡(Pyroptosis)是一种炎症性程序性细胞死亡,主要通过炎症小体依赖的经典途径(激活Caspase-1)或非经典途径(如Caspase-4/5/11)剪切Gasdermin家族蛋白(如GSDMD),剪切后的GSDMD-N端寡聚化并在细胞膜上形成孔道,导致细胞渗透压失衡、破裂,并释放IL-1β等促炎因子[1~2]。
其形态学特征主要表现为细胞膜上Gasdermin蛋白介导的孔洞形成,导致细胞肿胀、膜破裂并释放内容物,同时伴随核浓缩(但无典型核碎裂)和细胞器膨胀,最终细胞膜完整性丧失并引发强烈的炎症反应[3~5]。
细胞焦亡与多种疾病密切相关,包括感染性疾病、自身免疫性疾病、心血管疾病和癌症等,过度或持续的细胞焦亡可通过放大炎症反应导致组织损伤,而靶向调控焦亡通路已成为相关疾病治疗的潜在策略[6~7]。
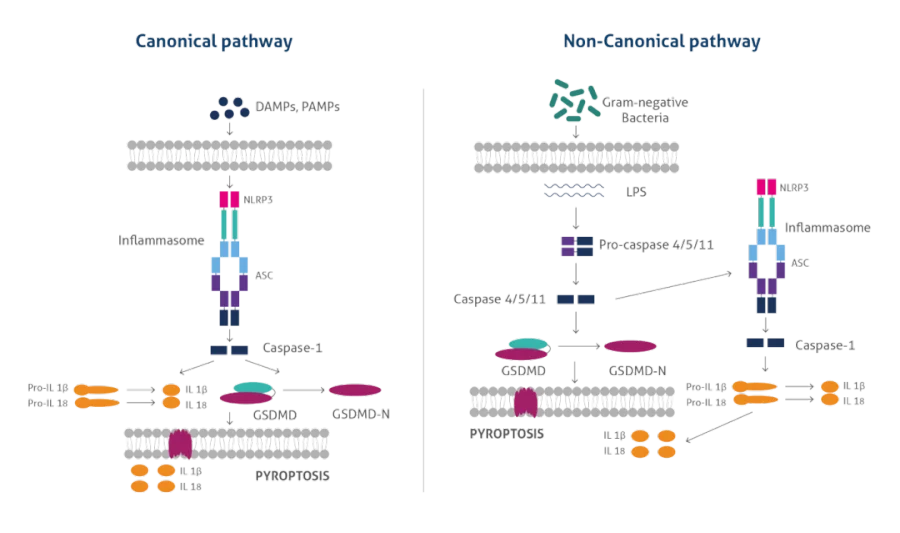
图1 细胞焦亡的经典和非经典途径
2、GSDMD、Caspase 1检测技巧解析
(上下滑动查看全部)
GSDMD(Gasdermin D)
GSDMD(消皮素D)是细胞焦亡的关键执行者,当细胞受到病原体感染或危险信号刺激时,炎症小体(如NLRP3、AIM2等)被激活,进而激活Caspase-1(经典途径)或Caspase-4/5/11(非经典途径)。活化的Caspase切割GSDMD,释放其N端结构域(GSDMD-N),该片段易位至细胞膜并寡聚化,形成孔道,导致细胞渗透压失衡、肿胀破裂,释放炎症因子和细胞内容物,引发强烈炎症反应[1~5]。
组织表达特异性:广泛表达于几乎所有类型的组织和细胞中。
分子量:GSDMD存在多种蛋白形式,包括约53 kDa的GSDMD前体、约31 kDa的剪切体GSDMD-N(活性形式)、约22 kDa的剪切体GSDMD-C等。
亚细胞定位:GSDMD以无活性的全酶形式(53 kDa)定位于细胞质,GSDMD-N端定位于细胞膜内侧(质膜)。
GSDMD检测Tips:
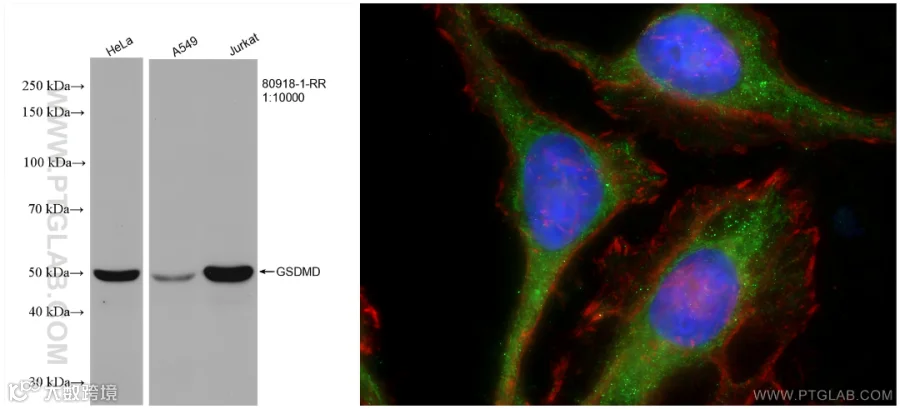
使用GSDMD抗体的WB(80918-1-RR)和IF(20770-1-AP)检测图
Caspase-1
Caspase-1(半胱天冬酶-1)是一种关键的炎症相关蛋白酶,属于半胱氨酸蛋白酶家族。当细胞感知病原体感染或危险信号时,Caspase-1被激活并切割Gasdermin D(GSDMD),释放其N端结构域(GSDMD-N),后者在细胞膜上形成孔道,导致细胞渗透性肿胀、内容物释放,最终引发细胞焦亡。同时,Caspase-1还剪切并激活IL-1β和IL-18前体,促进这些炎性因子的分泌,放大免疫应答[9]。
组织表达特异性:广泛表达于几乎所有类型的组织和细胞中。
分子量:酶原形式(Pro-caspase-1)的分子量为45 kDa,激活后会自身切割形成两个亚基:p20亚基(约20 kDa,与炎症小体复合物结合的关键片段)和p10亚基(约10 kDa)。
亚细胞定位:无活性的酶原(pro-caspase-1)形式主要位于细胞质。
Caspase 1检测Tips:
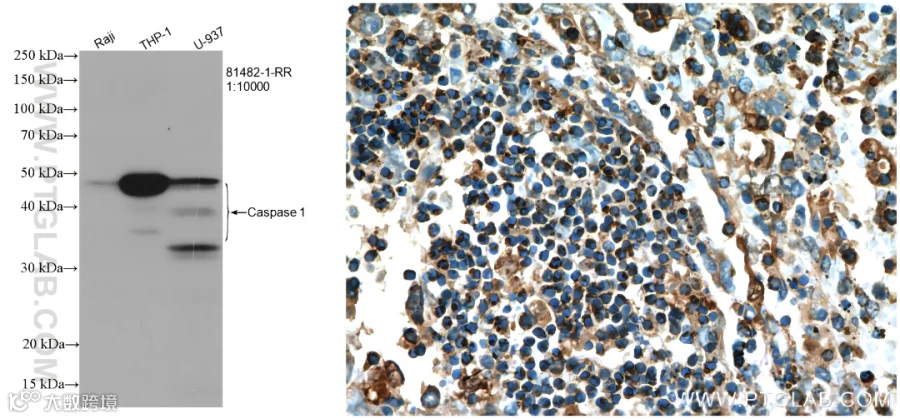
使用Caspase-1抗体的WB(81482-1-RR)和IHC(22915-1-AP)检测图
3、焦亡相关抗体推荐
焦亡主要由Gasdermin家族蛋白(如GSDMD)介导。关键靶点包括NLRP3炎症小体、Caspase-1/4/5/11等,它们激活后切割GSDMD,释放其N端结构域在膜上形成孔洞,导致细胞肿胀破裂并释放IL-1β、IL-18等炎症因子。
更多产品请登录官网查看!(点击蓝字可直达)
1、什么是铜死亡?
(上下滑动查看全部)
铜死亡代表一种独特的细胞死亡方式,其过程涉及对铜离子的依赖、脂酰化蛋白的积累以及铁硫簇蛋白的减少,导致细胞发生不同于氧化应激相关的细胞死亡(如凋亡、铁凋亡和坏死)的新型调节性细胞死亡。
在正常情况(铜稳态)下:铜离子在细胞内保持稳态,主要依赖于三个铜转运蛋白SLC31A1、ATP7A以及ATP7B。SLC31A1负责摄入铜,并依赖ATP7A/B负责转出铜,从而维持细胞内铜离子的动态平衡;
在非正常情况下:铜离子在依赖线粒体呼吸的细胞中发生累积,并通过与硫辛酰化DLAT结合,致使其发生异常的寡聚,形成不溶性DLAT,诱导细胞死亡;另一方面,FDX1能够将铜离子还原成更加具有毒性的一价铜离子,抑制铁硫簇蛋白的合成,从而诱导细胞死亡[10]。

图2 铜死亡[12]
2、SLC31A1、FDX1检测技巧解析
(上下滑动查看全部)
SLC31A1
SLC31A1是细胞膜上的铜转运蛋白,负责铜离子的跨膜运输。其功能缺失会导致线粒体铜耗竭,进而影响能量代谢。
组织表达特异性:广泛表达,多种肿瘤组织中表达水平上调。
分子量:计算分子量为21 kDa,实际检测分子量为21-25 kDa,35-40 kDa,60-70 kDa。
亚细胞定位:细胞膜(多次跨膜蛋白)。
SLC31A1检测Tips:
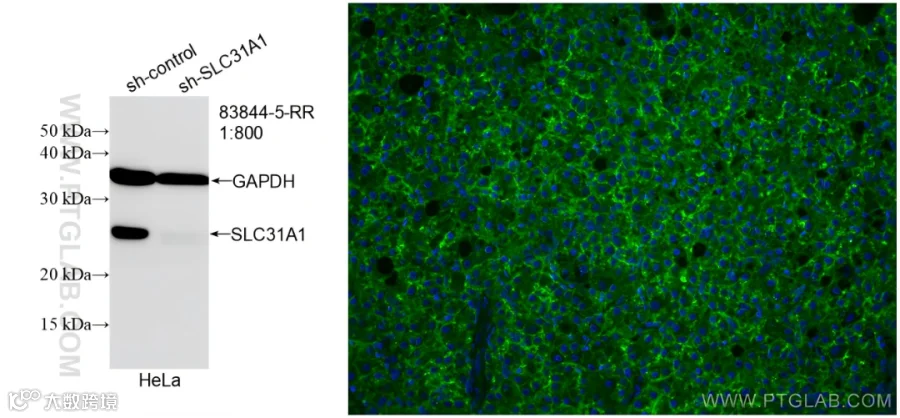
使用SLC31A1抗体的KO验证图(83844-5-RR)和IF(27499-1-AP)检测图
FDX1
FDX1主要参与线粒体内的电子转移过程,是铜死亡过程中的核心靶标之一。它在铜离子载体(如elesclomol)诱导的铜死亡中起关键作用。FDX1能够将Cu²⁺还原为Cu⁺,这种还原形式的铜具有更高的细胞毒性。此外,FDX1还通过促进DLAT的硫辛酰化,使DLAT与Cu⁺结合并寡聚化,导致蛋白质毒性应激和细胞死亡。
组织表达特异性:正常组织中表达,脑组织中高表达,在部分肿瘤组织中表达水平降低。
分子量:14-19 kDa。
亚细胞定位:主要定位于线粒体,内质网和细胞质中也有所表达。
FDX1检测Tips:
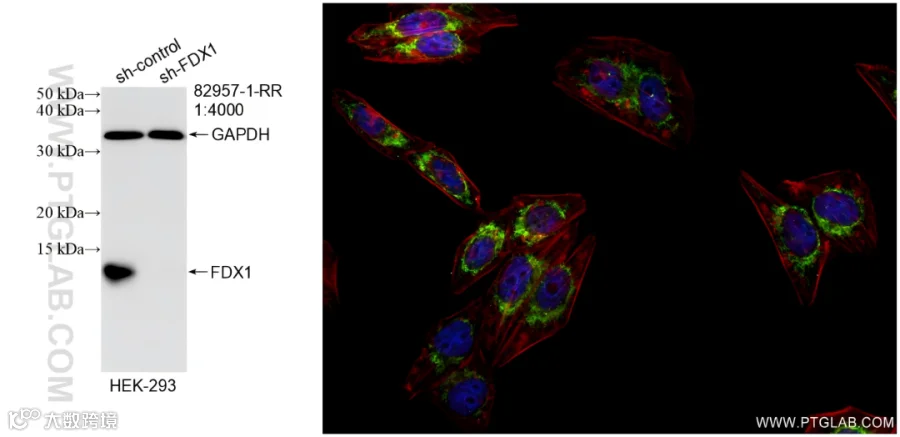
使用FDX1抗体(82957-1-RR)的KO验证和IF检测图
3、铜死亡相关抗体推荐
铜死亡是一种铜依赖性细胞死亡方式,主要由过量铜离子与线粒体三羧酸循环(TCA)中的脂酰化蛋白DLAT结合,导致蛋白聚集和毒性应激,引发细胞死亡。关键靶点包括FDX1(铁氧还蛋白1)、LIAS(脂酰转移酶)及DLAT(二氢硫辛酰胺转乙酰酶)。
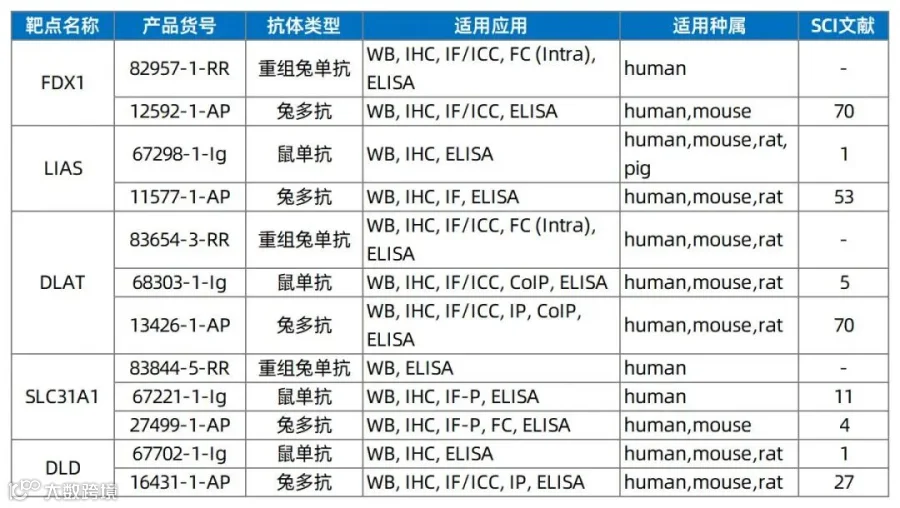
更多产品请登录官网查看!(点击蓝字可直达)
1、什么是双硫死亡?
(上下滑动查看全部)
双硫死亡 (Disulfidptosis)是由于细胞内二硫化物(如胱氨酸)异常积累而引发的一种程序性细胞死亡形式。当SLC7A11高表达的癌细胞,在葡萄糖饥饿条件下,NADPH合成减少,从而抑制了胱氨酸向半胱氨酸的转化过程。导致胱氨酸的大量积累。大量二硫化物的积累,从而引发了细胞死亡,即双硫死亡[13,14]。

图3 双硫死亡与胞内二硫化物氧化还原过程的分子机制[13]
2、SLC7A11/xCT、GLUT1检测技巧解析
(上下滑动查看全部)
SLC7A11/xCT
组织表达特异性:SLC7A11在肝癌、肺癌、胰腺癌、恶性胶质瘤中表达均增高。
分子量:SLC7A11的预估分子量约为55 kDa,主流分子量在35-40 kDa,可能为截短形式。
亚细胞定位:通常定位于细胞膜。
SLC7A11/xCT检测Tips:
1. WB优化建议
(1)样本处理:SLC7A11是跨膜蛋白,建议上样前避免常规煮沸(100℃),可尝试不煮样或37℃煮样,以防止蛋白聚集。
(2)分子量注意:理论分子量约55 kDa,但实际检测中常出现35-40 kDa 条带(可能为截短形式)。
2. IF检测优化
(1)固定与通透:SLC7A11为多次跨膜蛋白,需先固定(如4% PFA)后通透,建议使用温和通透剂(如Digitonin、Saponin 或低浓度的Triton X-100),避免过度破坏膜结构。
(2)抗体选择:优先推荐IF验证通过的抗体。
3. 其他建议
(1)阳性对照:使用高表达SLC7A11的细胞(如细胞系Hela)作为对照。
(2)刺激处理:可尝试erastin处理(抑制xCT功能)。
(3)推荐使用膜蛋白提取试剂盒进行样本制备(货号:PK10015)。
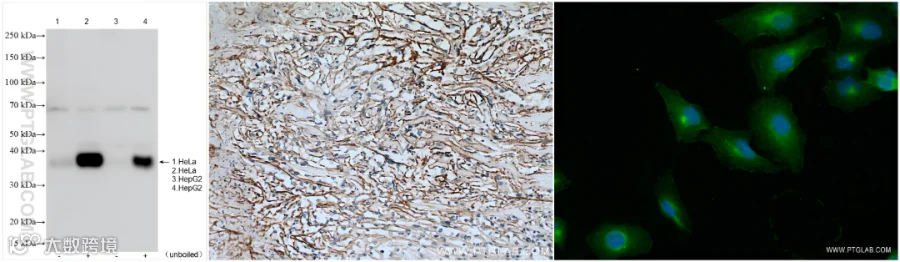
使用SLC7A11抗体的WB(26864-1-AP)、IHC(26864-1-AP)和IF(82115-2-RR)检测图
GLUT1
GLUT1,也被称为SLC2A1,是一种普遍表达的葡萄糖转运蛋白,在大多数细胞类型中负责葡萄糖摄取的基础水平,人红细胞表达GLUT1的水平最高。缺陷是导致1型和2型GLUT1缺乏症的原因。据报道,GLUT1的高表达是青少年血管瘤的可靠免疫组织化学标志物。
组织表达特异性:在红细胞中检测到。在许多人体组织中以不同水平表达。
分子量:主流分子量是50kDa。由于该蛋白是多次跨膜蛋白,且存在糖基化和磷酸化修饰,实际检测MW在43 kDa至55 kDa,且可能出现多条带。
亚细胞定位:通常定位于细胞膜。
GLUT1实验检测技巧
1. WB优化建议
(1)样本处理:GLUT1是跨膜蛋白,常规煮沸可能导致蛋白聚集,建议尝试不煮样或37℃煮样。
(2)GLUT1是多次跨膜蛋白,且存在糖基化和磷酸化修饰,在43 -55 kDa之间可能检出多条带。
2. IF检测优化
(1)固定与通透:GLUT1为多次跨膜蛋白,需先固定(如4% PFA)后通透,建议使用温和通透剂(如Digitonin、Saponin 或低浓度的Triton X-100),避免过度破坏膜结构。
(2)抗体选择:优先推荐IF验证通过的抗体。
3.推荐使用膜蛋白提取试剂盒进行样本制备(货号:PK10015)。
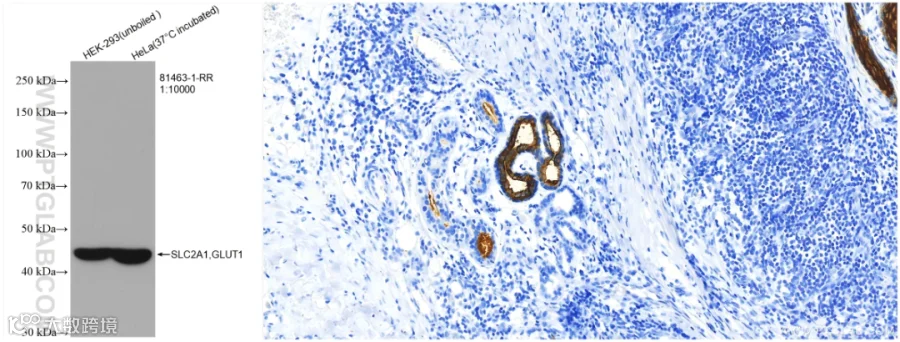
使用GLUT1抗体(81463-1-RR)的WB和IHC检测图
3、双硫死亡相关抗体推荐
双硫死亡机制涉及NADPH耗竭导致二硫化物应激,使肌动蛋白(如SLC7A11、ACTB)发生异常二硫键交联,破坏细胞骨架稳定性,最终引发细胞死亡。
更多产品请登录官网查看!(点击蓝字可直达)
1、什么是失巢凋亡?
(上下滑动查看全部)
失巢凋亡(Anoikis,希腊语为“无家可归者”)是一种特殊的细胞程序死亡,它是由细胞与细胞外基质或相邻细胞脱离接触而诱发的。当正常细胞失去对细胞外基质的粘附时,就会发生细胞凋亡,因为机体阻止了细胞之间的错误接触。
然而,肿瘤细胞具有转移特性,它失去对细胞外基质的粘附后,会产生耐药性,也称为失巢凋亡抗性(anoikis resistance)。在不粘附到细胞外基质的情况下也可以存活,并且可以通过血液和淋巴液流向其他组织和器官进行粘附和转移[15]。

图4 细胞与细胞外机制粘附丧失的细胞进程[15]
2、FAK、PI3K p110 Beta检测技巧解析
(上下滑动查看全部)
FAK
FAK(Focal Adhesion Kinase,黏着斑激酶)是一种非受体酪氨酸激酶,在细胞信号转导中起着关键作用。它广泛存在于真核细胞中,参与细胞黏附、迁移、增殖、存活以及细胞周期调控等多个生物学过程。FAK的异常激活与肿瘤细胞的侵袭、转移、增殖和凋亡抑制密切相关。
组织表达特异性:肿瘤中的高表达和在正常组织中的低表达。
分子量:100-120kDa和40-60kDa。
亚细胞定位:主要位于细胞质,细胞核和细胞外基质也有表达。
FAK检测Tips:
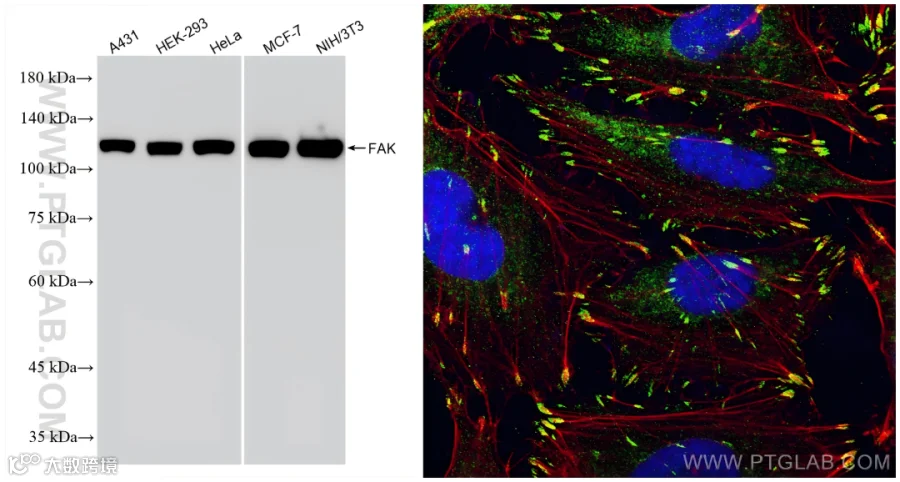
使用FAK抗体的WB(81334-5-RR)和IF(12636-1-AP)检测图
PI3K p110 Beta
PI3 Kinase p110 Beta(磷脂酰肌醇3激酶p110β)是PI3K(磷脂酰肌醇3激酶)家族中的一个催化亚基,属于Class IA PI3Ks。PI3Ks在细胞信号传导中起着关键作用,通过生成磷脂酰肌醇-3,4,5-三磷酸(PIP3)来调节多种细胞过程,包括细胞生长、存活、迁移和代谢。PI3K是FAK激活的蛋白之一,它反过来招募并激活其下游靶蛋白激酶B (PKB/Akt)[16]。
组织表达特异性:p110β在多种组织和细胞类型中广泛表达。
分子量:110-130 kDa。
亚细胞定位:位于细胞质、细胞核。
PI3 Kinase p110 Beta检测Tips
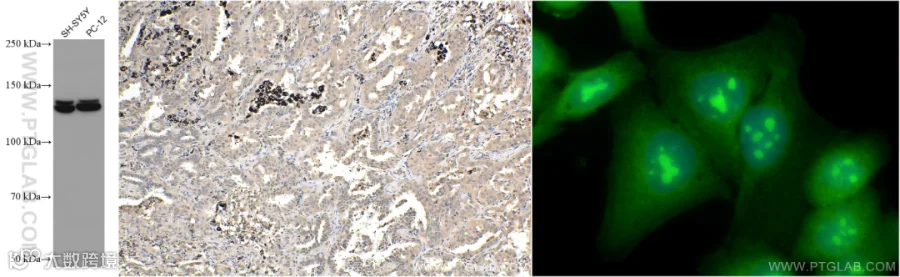
使用PI3 Kinase p110 Beta抗体的WB(67121-1-lg)、IHC和IF检测图(20584-1-AP)
3、失巢凋亡相关抗体推荐
失巢凋亡关键靶点包括FAK/PI3K通路抑制、Bcl-2家族失衡(如Bim上调)、caspase级联反应(如caspase-3/8)等。该机制可阻止细胞异常迁移,若失调则促进肿瘤转移。EMT过程可通过激活ERK/MAPK或NF-κB通路来协助癌细胞抵抗失巢凋亡。
更多产品请登录官网查看!(点击蓝字可直达)
1、什么是泛凋亡?
(上下滑动查看全部)
PANoptosis(泛凋亡)是一种新定义的PCD,于2019年首次提出,同时涉及焦亡(P)、凋亡(A)和坏死(N)的关键分子。与其他形式类似,它可以被多种触发因素刺激,如病毒、细菌和细胞因子。与细胞凋亡相反,PANoptosis是一种类似于焦亡和坏死的炎症过程,导致细胞膜破坏,细胞因子和损伤相关分子模式(DAMPs)的释放[17]。
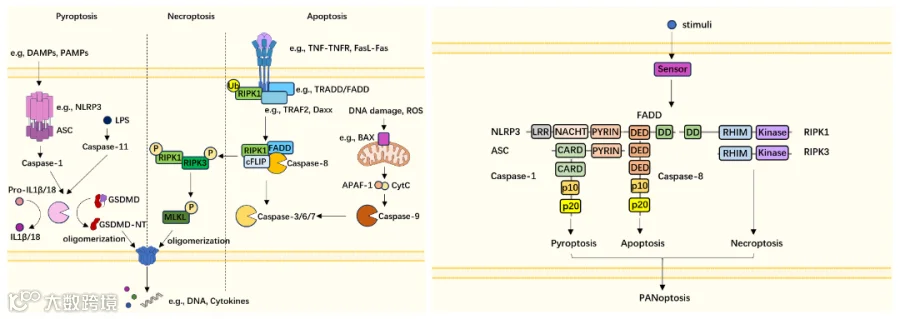
图5 凋亡、焦亡、坏死性凋亡的分子机制(左);泛凋亡复合体(PANoptosome)组分(右)[17]
2、Caspase 8检测技巧解析
(上下滑动查看全部)
Caspase 8
Caspase-8是调控泛凋亡(panoptosis)的关键分子,作为细胞凋亡的启动酶,它既能激活经典凋亡通路(切割caspase-3),也可通过抑制RIPK3/MLKL通路阻断坏死性凋亡(necroptosis)。在泛凋亡复合体(panoptosome)中,Caspase-8与炎症小体组分(如NLRP3)协同作用,整合凋亡、焦亡和坏死性凋亡信号,触发混合型细胞死亡并释放大量炎症因子,参与抗感染和肿瘤免疫调控。
组织表达特异性:在多种组织中广泛表达。
分子量:约55 kDa。
亚细胞定位:位于细胞质和细胞核。
Caspase-8检测Tips:
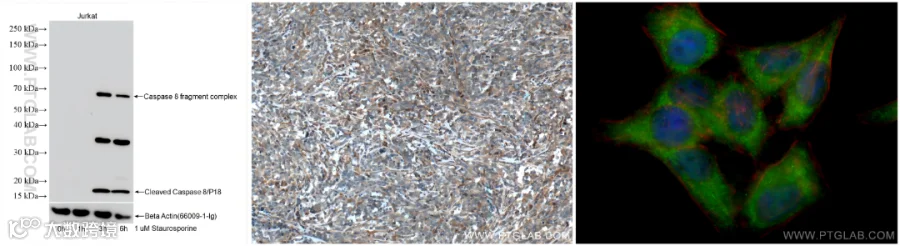
使用Caspase-8抗体的WB(85171-3-RR)、IHC和IF检测图(13423-1-AP)
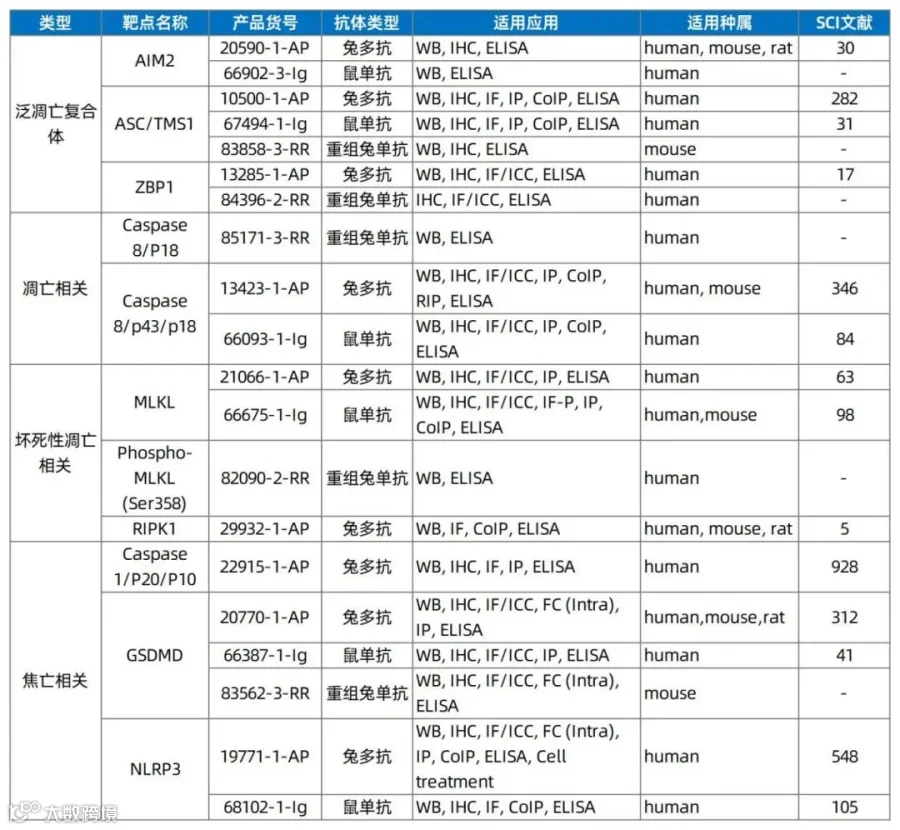
3、泛凋亡相关抗体推荐
泛凋亡核心靶点包括PANoptosome复合体(含ZBP1、RIPK1、caspase-8等),通过激活caspase-3/8(凋亡)、GSDME(焦亡)和MLKL(坏死性凋亡)等效应分子,协同触发炎症性细胞死亡。
更多产品请登录官网查看!(点击蓝字可直达)
以上是十种细胞死亡形式,及其热门靶标检测技巧,希望能够给从事相关研究的老师们一些帮助!
下期预告:我们将继续解析细胞增殖研究及关键靶点检测技巧。欢迎持续关注,下期见~
参考文献
[1] Shi, Jianjin et al. “Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death.” Nature vol. 526,7575 (2015): 660-5.
[2] Kayagaki, Nobuhiko et al. “Caspase-11 cleaves gasdermin D for non-canonical inflammasome signalling.” Nature vol. 526,7575 (2015): 666-71.
[3] Liu, Xing et al. “Inflammasome-activated gasdermin D causes pyroptosis by forming membrane pores.” Nature vol. 535,7610 (2016): 153-8.
[4] Shi, Jianjin et al. “Pyroptosis: Gasdermin-Mediated Programmed Necrotic Cell Death.” Trends in biochemical sciences vol. 42,4 (2017): 245-254.
[5] Galluzzi, Lorenzo et al. “Molecular mechanisms of cell death: recommendations of the Nomenclature Committee on Cell Death 2018.” Cell death and differentiation vol. 25,3 (2018): 486-541.
[6] Jorgensen, Ine, and Edward A Miao. “Pyroptotic cell death defends against intracellular pathogens.” Immunological reviews vol. 265,1 (2015): 130-42.
[7] You, Ruixuan et al. “Pyroptosis and Its Role in Autoimmune Disease: A Potential Therapeutic Target.” Frontiers in immunology vol. 13 841732. 25 May. 2022,
[8] Zhang, Bao-Li et al. “Inhibition of GSDMD activation by Z-LLSD-FMK or Z-YVAD-FMK reduces vascular inflammation and atherosclerotic lesion development in ApoE-/- mice.” Frontiers in pharmacology vol. 14 (2023): 1184588.
[9] Li, Yang, and Qianzhou Jiang. “Uncoupled pyroptosis and IL-1β secretion downstream of inflammasome signaling.” Frontiers in immunology vol. 14 (2023): 1128358.
[10] Tsvetkov P, Coy S, Petrova B, et al. Copper induces cell death by targeting lipoylated TCA cycle proteins. Science. 2022 Mar 18;375(6586):1254-1261.
[11] Petris MJ. The SLC31 (Ctr) copper transporter family. Pflugers Arch. 2004 Feb;447(5):752-5.
[12] Tsui KH, Hsiao JH, Lin LT, et al. The Cross-Communication of Cuproptosis and Regulated Cell Death in Human Pathophysiology. Int J Biol Sci. 2024 Jan 1;20(1):218-230.
[13] Liu, Xiaoguang et al. Cystine transporter regulation of pentose phosphate pathway dependency and disulfide stress exposes a targetable metabolic vulnerability in cancer. Nature cell biology vol. 22,4 (2020): 476-486.
[14] Liu, Xiaoguang et al. Actin cytoskeleton vulnerability to disulfide stress mediates disulfidptosis. Nature cell biology vol. 25,3 (2023): 404-414.
[15] Han YH, Wang Y, Lee SJ, et al. Regulation of anoikis by extrinsic death receptor pathways. Cell Commun Signal. 2023 Sep 4;21(1):227.
[16] Paoli P, Giannoni E, Chiarugi P. Anoikis molecular pathways and its role in cancer progression. Biochim Biophys Acta. 2013 Dec;1833(12):3481-3498.
[17] Chen S, Jiang J, Li T, Huang L. PANoptosis: Mechanism and Role in Pulmonary Diseases. Int J Mol Sci. 2023 Oct 19;24(20):15343.
(上下滑动查看全部)


扫码添加云南总代理云南泽浩客服微信






往期文章推荐
Proteintech 抗体50%补贴、重组流抗首单5折、买1赠1、6倍积分(进行中)
Proteintech FlexAble抗体标记试剂盒低至7折(进行中)
