

神经元由胞体、细而长的轴突、粗而短且有许多分支的树突三部分组成。由于不同的代谢需求,这些区域中的线粒体分布并不均匀。突触、生长锥、轴突分支部位和郎飞氏结代谢活跃且对ATP需求巨大,因此往往分布着更多的线粒体。
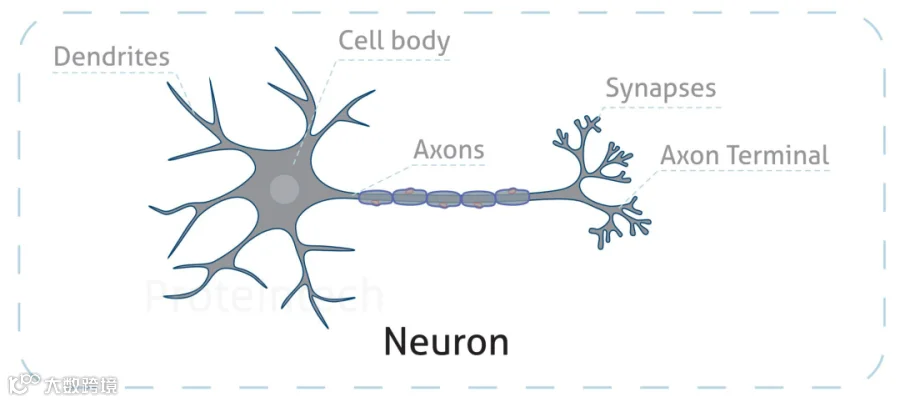
图1:神经元结构示意图(图源:Proteintech)
神经元是极化细胞,细胞骨架为神经元提供支撑和骨干,并允许细胞器在神经元内高效运输和锚定。神经元细胞骨架主要由微管(Microtubules)、肌动蛋白丝(Actin Filaments)和神经丝(Neurofilaments)组成。虽然线粒体的生物合成可以在局部发生,如在轴突内,但研究者们认为大多数新的线粒体是在胞体中生成的。功能失调的线粒体也会返回胞体,由自噬-溶酶体系统降解。
如果将微管比作是神经元细胞中的“具有双向车道的高速公路”,细胞器为“货物”,动力蛋白(Dynein)或驱动蛋白(Kinesin)则作为“卡车”载着这些货物在这条高速公路上来回行驶,将货物传递到ATP缺乏和细胞内Ca2+水平升高的区域。如线粒体在突触中非常活跃是为了产生ATP并缓冲Ca2+以维持神经元功能。
下面我们分别介绍线粒体在神经元细胞内外的运输机制。
从细胞体到远端轴突和树突区域的长距离线粒体运输依赖于神经元微管的极性和组织结构。微管由α-微管蛋白和β-微管蛋白聚合而成,并以极化方式排列,具有正负端。
线粒体必须与马达蛋白(Motor Protein,又称运动蛋白)偶联才能实现极化运输,其中驱动蛋白(Kinesin)介导指向远端的顺向运输,动力蛋白(Dynein)促进向胞体的逆向运输,肌球蛋白(Myosin)介导线粒体在微丝中短距离内的运动和锚定。这些马达蛋白的驱动力来自线粒体呼吸产生的ATP。由于树突的微管在近端区域表现出混合极性,驱动蛋白和动力蛋白可以根据微管极性驱动树突中的货物向前或向后移动。
线粒体通过两种方式与马达蛋白结合:第一种是线粒体直接与马达蛋白结合,第二种是与线粒体受体-衔接蛋白-马达蛋白的结合。这些马达蛋白-衔接蛋白-受体复合物确保了线粒体的靶向运输和对其移动性的精确调控。
2.1 线粒体在轴突中的运输
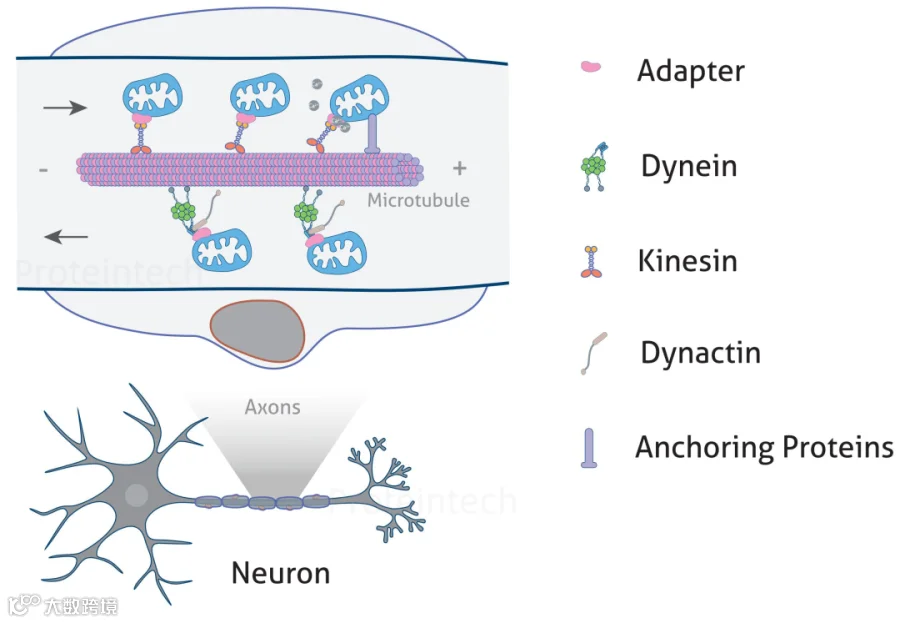
图2:线粒体在轴突中的运输(图源:Proteintech)
驱动蛋白在人类和小鼠中是由45种不同基因编码的驱动蛋白超家族蛋白(也称为KIFs)。KIF5的成员在神经元线粒体的顺向运输中起着关键作用。哺乳动物有三种KIF5运动亚型(即KIF5A、KIF5B和KIF5C)。KIF5B在大多数细胞类型中表达,而KIF5A和KIF5C仅存在于神经元。另外还有KIF1B和KLP6也是介导神经元线粒体运输的主要驱动蛋白。
在轴突中,线粒体通过衔接蛋白(Adapter)募集马达蛋白。不同的马达蛋白可能需要不同的衔接蛋白配合。在线粒体外膜上,线粒体Rho GTPase蛋白(MIRO1和MIRO2)通过TRAK衔接蛋白或MTX衔接蛋白募集马达蛋白。MIRO蛋白的C端包含一个跨膜结构域和两个GTPase 结构域,一个位于N端,另一个靠近C端。GTPase结构域可以影响线粒体的分布:当GDP结合时,MIRO不会募集衔接蛋白和马达蛋白。驱动蛋白KIF5与MIRO-TRAK1复合物结合将线粒体顺向运送到轴突末端,动力蛋白与MIRO-TRAK2复合物结合介导线粒体的逆向运输或短距离运输。TRAK不仅参与线粒体运输,还参与其他细胞器(包括内体)的运输。
Syntabulin是另一种KIF5的衔接蛋白,其C端跨膜结构域可实现线粒体靶向。与MIRO(通过与TRAK结合间接连接到KIF5驱动蛋白)不同,Syntabulin直接与KIF5结构域相互作用,从而介导KIF5与线粒体的结合。
动力蛋白是一个包含11个亚基的大型蛋白质复合物。它包括两条催化动力蛋白重链(DHC)、多条动力蛋白中间链(DIC)、动力蛋白轻链中间链(DLIC)和动力蛋白轻链(DLC)。动力蛋白-动力蛋白激活蛋白复合物如何连接线粒体进行逆向运输的机制尚不明确。一种假设是动力蛋白通过其轻链和中间链与细胞器膜相关蛋白连接。如DYNLT1 是一种动力蛋白轻链蛋白,它与线粒体外膜蛋白电压依赖性阴离子选择性通道蛋白(VDAC1)结合形成复合物进行逆向运输。
2.2 线粒体在轴突末端的运输
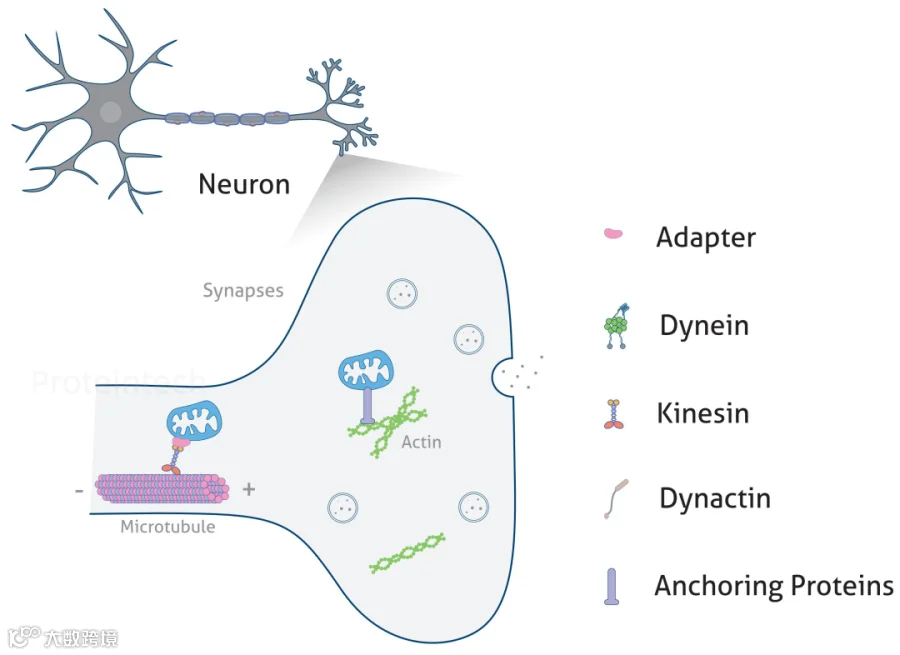 图3:线粒体在轴突末端的运输(图源:Proteintech)
图3:线粒体在轴突末端的运输(图源:Proteintech)
线粒体在轴突末端的主要功能是提供能量,并支持神经递质的合成和释放。线粒体的长距离运输是由基于微管的驱动蛋白和动力蛋白驱动的,但神经末梢和生长锥中的短距离运动很可能是由肌球蛋白介导的。尽管线粒体与肌球蛋白的关联以及肌球蛋白驱动的线粒体运输尚未在神经元中直接证实,但已证明与肌球蛋白V(MYO5)相关的货物沿轴突移动的速度明显低于大多数轴突细胞器的速度,与运输线粒体的速度相似。因此,肌球蛋白很可能驱动在肌动蛋白丝富集区域中的线粒体短距离运动。
最近一项使用果蝇神经元和RNAi的研究表明,肌球蛋白V(MYO5)的缺失增加了线粒体在两个方向上的移动速度,而降低肌球蛋白VI(MYO6)的表达则选择性地增加了轴突中的逆向线粒体运输。这些结果表明肌球蛋白V和VI可能与基于微管的马达蛋白竞争。或者,MYO5/MYO6和动力蛋白竞争与线粒体的结合,导致它们在逆向运动过程中从微管轨道移位到肌动蛋白丝上。
肌球蛋白XIX(MYO19)也是一种新发现的线粒体相关肌球蛋白,参与线粒体基于肌动蛋白丝运输。研究表明在A549细胞中表达绿色荧光蛋白(GFP)-MYO19融合蛋白可显著增强基于肌动蛋白丝的线粒体运动。MYO19通过其尾部结构域靶向线粒体,且在包括神经元在内的多种细胞中广泛表达,因此MYO19也可能参与沿神经元突起和突触末梢的线粒体运输。
2.3 线粒体的锚定
线粒体锚定是在特别依赖能量产生(即ATP)和 Ca2+丰富的区域中需要维持一定数量的静止线粒体。线粒体相关蛋白 Syntaphilin (SNPH)是一种神经元特异性蛋白。Syntaphilin通过其N端微管结合域直接与微管相互作用,而其C端尾部插入线粒体外膜。高浓度的Ca2+充当“停止”信号,Ca2+与MIRO/TRAK复合物结合,马达蛋白与线粒体结合减弱,Syntaphilin则将线粒体锚定在微管上,线粒体运输停止并释放能量。
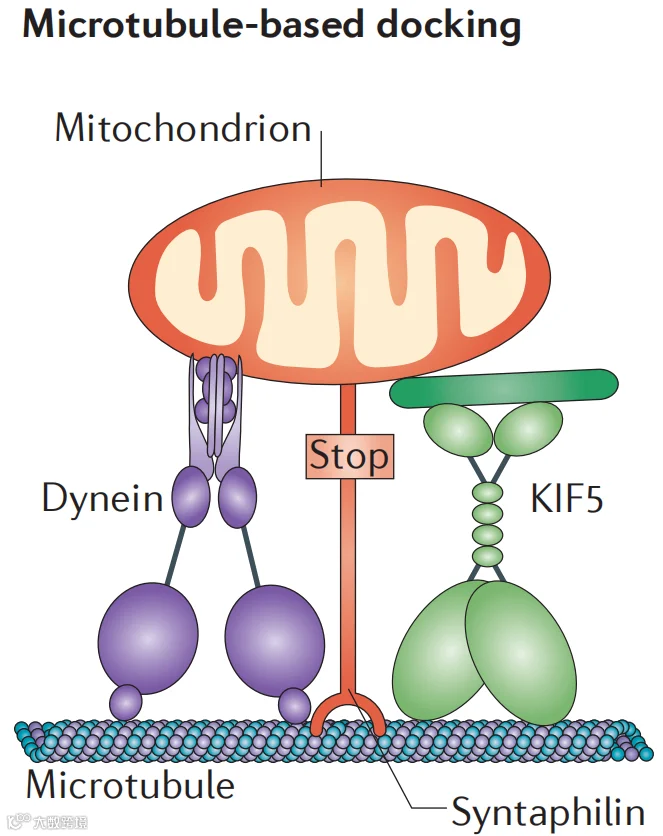
图4:线粒体在微管的锚定(图源:参考文献2)
线粒体在肌动蛋白丝的短距离运输中所涉及的锚定系统尚未确定。研究发现,神经生长因子(NGF)可以影响线粒体与肌动蛋白之间的相互作用,从而调节线粒体的迁移率。WASP家族富含脯氨酸同源蛋白1(WAVE1)调节肌动蛋白聚合,对于线粒体向树突棘的运输至关重要。
正常的神经元稳态不仅依赖于细胞内的线粒体运输,也依赖于细胞间的线粒体交换。例如,星形胶质细胞可以将健康的线粒体转移到受损的神经元,并提供神经保护和神经修复。
线粒体跨细胞转移的多种机制已被鉴定,包括隧道纳米管(TNT:tunnelling nanotubes)、细胞外囊泡(EV:extracellular vesicles)和间隙连接通道(GJC:gap junction channels)。还有一些其他的线粒体转移模式,例如细胞融合(cell fusion)。
1) TNT是连接细胞的瞬时纤维膜,由细胞膜、肌动蛋白、肌球蛋白和微管蛋白组成。TNT主要存在于应激细胞和非应激细胞之间,能够运输多种细胞成分。它是连接两个或多个细胞的膜突起,能形成瞬时胞质桥,从而实现了货物的双向运输。线粒体是通过TNT进行运输的重要实体。间充质细胞(MSC)可以形成TNT并将线粒体和其他成分转移到靶细胞。
2) EVs包括外泌体、微泡和凋亡小体,是细胞分泌的纳米级双层结构囊泡,可携带各种脂质、蛋白质、RNA、miRNA和线粒体。研究发现星形胶质细胞释放线粒体颗粒,然后部分被受损神经元吸收,从而促进神经元存活和树突再生。
3) GJC是两个相邻细胞之间最直接的物质交换通道,由细胞各自的半通道(HC:hemichannels)对接而成。间隙连接通道允许分子量高达1 kDa的分子被动扩散,并运输营养物质、代谢物、第二信使、阳离子、阴离子和线粒体等。
4) 细胞融合是线粒体非经典转移方式。细胞融合可以在体内自发发生,也可以人工诱导。自发细胞融合,主要是干细胞与体细胞的融合。这种融合有助于体细胞的重编程和组织的再生。
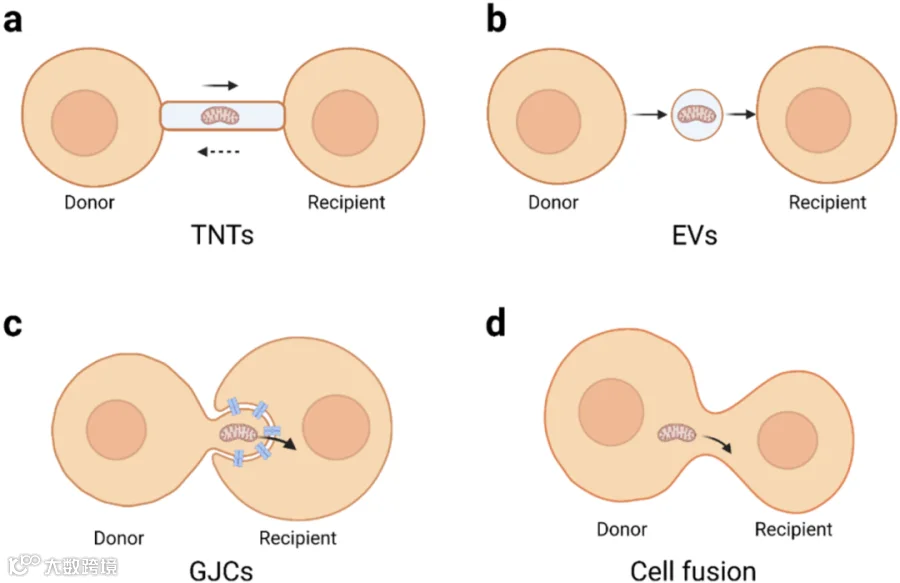
图5:细胞间线粒体转移模式(图源:参考文献4)
在神经元发育过程中,大部分线粒体会通过移动来供能,但在成熟的神经元中,静止的线粒体占了三分之二以上,它们充当着“发电站”的角色。线粒体运输障碍会导致神经元功能受损,并与多种神经系统疾病相关。例如,在阿尔茨海默病中,tau过表达可以选择性地抑制驱动蛋白前向运输,并导致线粒体在细胞体中积累,突触末端线粒体减少;肌萎缩侧索硬化症患者(ALS)和ALS小鼠模型表现出轴突线粒体运输受损等。
目前,线粒体在神经元中的运输的研究正处于快速上升阶段,“神经元如何精确控制基于微管和肌动蛋白丝的运输之间的转换”,“线粒体在肌动蛋白丝的短距离运输中所涉及的锚定系统”以及“肌动蛋白或肌球蛋白如何连接线粒体并驱动其运输”等等机理仍未被研究透彻。有待研究者们深入探究。
Proteintech线粒体运输研究相关抗体非常齐全,适用于WB、IP、IF、IHC、FC等多种实验场景。小P还为大家准备了线粒体动力学全新海报供学习参考,点击蓝字可领取哦~
(↓上下滑动以查看全部↓)
参考文献
(↓上下滑动以查看全部↓)
1. Lu D, et al. Mitochondrial transport in neurons and evidence for its involvement in acute neurological disorders. Front Neurosci. 2023 Oct 12;17:1268883.
2. Sheng ZH, Cai Q. Mitochondrial transport in neurons: impact on synaptic homeostasis and neurodegeneration. Nat Rev Neurosci. 2012 Jan 5;13(2):77-93.
3. Quintero OA, et al. Human Myo19 is a novel myosin that associates with mitochondria. Curr Biol. 2009 Dec 15;19(23):2008-13.
4. Liu Z, et al. Mitochondrial transfer/transplantation: an emerging therapeutic approach for multiple diseases. Cell Biosci. 2022 May 19;12(1):66.
5. Shanmughapriya S, Langford D, Natarajaseenivasan K. Inter and Intracellular mitochondrial trafficking in health and disease. Ageing Res Rev. 2020 Sep;62:101128.
6. Luchetti F, et al. Tunneling nanotubes and mesenchymal stem cells: New insights into the role of melatonin in neuronal recovery. J Pineal Res. 2022 Aug;73(1):e12800.


扫码添加云南总代理云南泽浩客服微信







往期文章推荐
