

泛读此文,请直接看花色字体快读,大概需1min;精读此文,大概需10min。

9月11日, 2018年拉斯克奖(“诺奖风向标”)结果公布,加州大学的Michael Grunstein和洛克菲勒大学的C. David Allis共同分享了今年的基础医学研究奖。获奖理由是发现并阐释了组蛋白化学修饰对基因表达的影响。其中,甲基化修饰是组蛋白化学修饰中一种比较常见的修饰,近几年的新生力量RNA甲基化更是把甲基化修饰家族带到了一个新的高度。限于篇幅,本文先分享RNA甲基化修饰的完整研究思路,后续再解读DNA、蛋白质的甲基化修饰研究思路。
最近几年,关于RNA甲基化的研究,热度逐渐抬高(见图1)。这个热点早在2015年就已经在《Nature》和《Cell》上发表了相关研究结果。
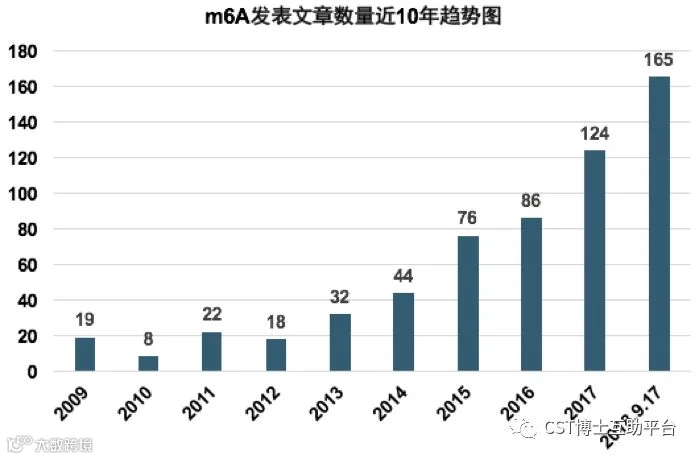
图1. m6A发表数量10年趋势图。2018年预计全年文章至少240篇,相比2017年增长94%,大约20篇/月的暴走节奏。2017年文章数量比2016年增长44%。
另外,2016年中标的国自然中m6A的项目数是5个,2017年有22个;2018年增加到了60个左右,增加到了近3倍,经费达到了近3000万。预计2019年中标的项目数会超过100项,经费级别达到5000万+。m6A已然成为中国科研界的“明日之星“。
图2. 百度搜索“RNA甲基化”结果有百万条
那么,RNA甲基化到底是什么?有哪些过人之处?有哪些研究工具可使用?有哪些研究套路可参考?
1
RNA甲基化是什么?有哪些过人之处?
已知RNA存在超过100种修饰,mRNA最常见的内部修饰包括N6-腺苷酸甲基化(m6A)、N1-腺苷酸甲基化(m1A)、胞嘧啶羟基化(m5C)等。RNA甲基化修饰约占所有RNA修饰的60%以上,而m6A是高等生物mRNA和lncRNAs上最为丰富的甲基化修饰。研究发现,microRNA、circRNA、rRNA、tRNA和snoRNA上都有m6A修饰的发生。
m6A英文全称是N6-methy-ladenosine,N6-甲基腺嘌呤,是一种动态可逆的修饰方式。如图3,m6A是指碱基A第6位N原子上的甲基,主要存在于mRNA的CDS区和3’UTR区。m6A在转录后调控中发挥重要作用,影响mRNA的稳定性,翻译效率,可变剪接,RNA的转运和衰变等。m6A RNA出现在多种重要的细胞生命过程中,意味着mRNA甲基化参与了多种生物学过程,一旦参与m6A修饰的酶出现异常,将会引起很多疾病,包括肿瘤、神经性疾病、代谢性疾病、胚胎发育迟缓等。
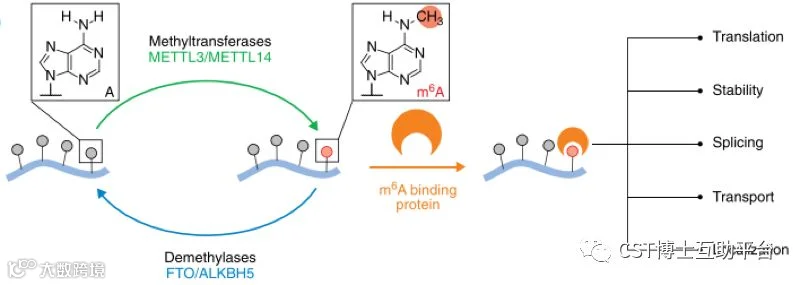
图3. m6A RNA修饰及介导的功能(图片来自网络)
类似于DNA甲基化、组蛋白甲基化修饰受一些编写酶(Writers)和去除酶(Erasers)(🔗1,🔗2)的调控,m6A修饰主要受去甲基化酶、甲基化酶和甲基化识别酶等调控,即由“编码器”(Writer)、“读码器”(Reader)及“消码器”(Eraser)协同起作用。
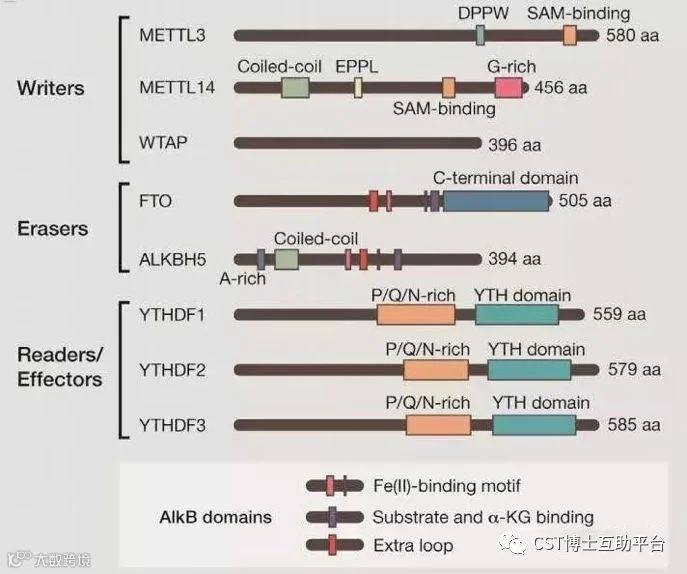
图4. 人m6A修饰酶的编码器、读码器和消码器的结构域
(图片来自Mihye Lee et at(2014)Cell,158:980)
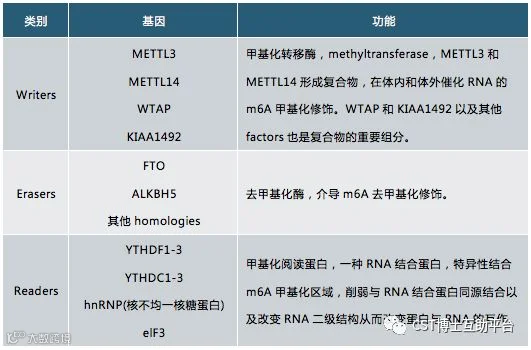
图5. 人m6A修饰酶的编码器、读码器和消码器的基因名及功能
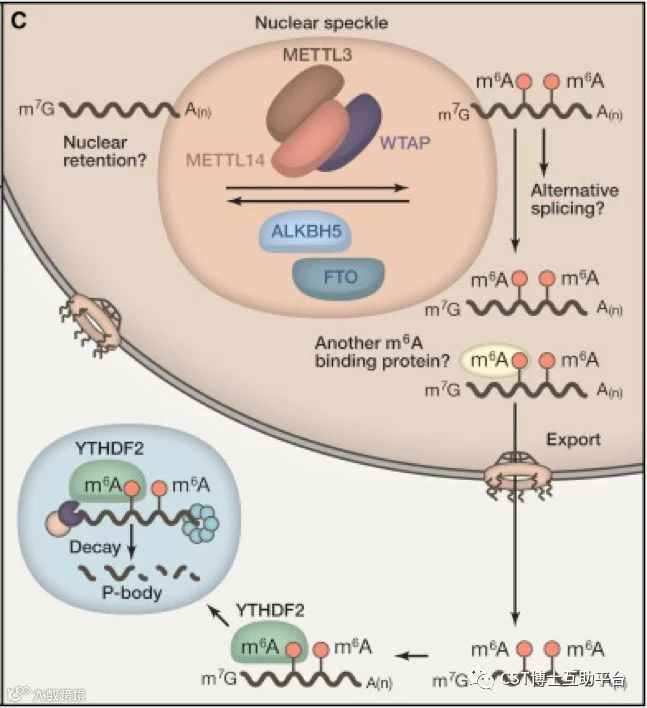
图6. m6A在mRNA上的细胞功能模型(图片来自Mihye Lee et at(2014)Cell,158:980)
2
【研究m6A有哪些工具可用?】
CST提供一整套RNA甲基化研究的科研工具,包括初筛Sampler Kit、抗体、siRNA、细胞因子、ChIP试剂盒、实验辅助试剂、激动剂和抑制剂等,具体见如下表。若需要用到相关产品,请联系CST授权一级代理基因有限公司订购。
类别 |
产品名称 |
货号 |
应用 |
反应性 |
文献数 |
|
Writers |
METTL3 (D2I6O) Rabbit mAb |
96391 |
WB IP |
H M R Mk |
||
METTL14 (D8K8W) Rabbit mAb |
51104 |
WB |
H M R Mk |
|||
WTAP Antibody |
56501 |
WB IP |
H M R Mk |
|||
Virilizer (D4N8B) Rabbit mAb |
88358 |
WB IP CHIP |
H M R Mk |
|||
Erasers |
FTO (D6Z8W) Rabbit mAb |
31687 |
WB IP |
H |
1 |
|
FTO (D2V1I) Rabbit mAb |
45980 |
WB |
H M R |
1 |
||
Readers |
YTHDC1 (D10) Antibody |
54737 |
WB |
H M R Mk |
||
YTHDC1 (E4I9E) Rabbit mAb |
77422 |
WB IP |
H M R Mk |
|||
hnRNP A2/B1 (2A2) Mouse mAb |
9304 |
WB F |
H Mk |
4 |
||
hnRNP C1/C2 (D6S3N) Rabbit mAb |
91327 |
WB |
H Mk |
1 |
||
hnRNP A0 (D8A3) XP® Rabbit mAb |
5545S |
WB IP IF F |
H M R Mk |
1 |
||
hnRNP A1 (D21H11) Rabbit mAb |
8443 |
WB IP IF F |
H M R Mk |
11 |
||
hnRNP K (R332) Antibody |
4675 |
WB IF F |
H M R Mk |
6 |
||
hnRNP E1 Antibody |
8534 |
WB IF |
H M R |
1 |
||
AUF1/hnRNP D (D6O4F) Rabbit mAb |
12382 |
WB IP IHC |
H |
|||
hnRNP Q/R (D18B2) Rabbit mAb |
8588 |
WB IP |
H M R Mk |
|||
hnRNP LL Antibody |
4783 |
WB IP |
H M R |
|||
hnRNP E2/PCBP2 (D1S5E) Rabbit mAb |
83017 |
WB |
H M R |
|||
hnRNP L Antibody |
37562 |
WB |
H M R Mk |
2 |
||
RBMX/hnRNP G (D7C2V) Rabbit mAb |
14794 |
WB IP |
H M R Mk |
|||
SignalSilence® hnRNP A1 siRNA I |
7668 |
|||||
SignalSilence® AUF1/hnRNP D siRNA I |
12763 |
|||||
eIF3A (D51F4) XP® Rabbit mAb |
3411 |
WB IP IHC IF |
H M R Mk |
7 |
||
eIF3C Antibody |
2068 |
WB |
H M Mk |
2 |
||
eIF3J (D21G7) XP® Rabbit mAb |
8161 |
WB IP IF |
H M R Mk |
|||
eIF3H (D9C1) XP® Rabbit mAb |
3413 |
WB IP IHC F |
H M R Mk |
6 |
||
eIF2α (D7D3) XP® Rabbit mAb |
5324 |
WB IP IHC |
H M R Mk |
90 |
||
Phospho-eIF2α (Ser51) (D9G8) XP® Rabbit mAb |
3398 |
WB IP IHC |
H M R Mk Dm |
155 |
||
eIF4H (D85F2) XP® Rabbit mAb |
3469 |
WB IP IF |
H M R Mk |
|||
eIF4B (1F5) Mouse mAb |
13088 |
WB IHC IF |
H |
1 |
||
Phospho-eIF4B (Ser406) Antibody |
5399 |
WB IP IF |
H M |
5 |
||
eIF4E (C46H6) Rabbit mAb |
2067 |
WB IP IHC |
H M R Mk |
27 |
||
eIF4G (C45A4) Rabbit mAb |
2469 |
WB IHC IF F |
H M R Mk |
14 |
||
Phospho-eIF4G (Ser1108) Antibody |
2441 |
WB IP IF |
H M R Hm Mk B |
35 |
||
eIF4G2/p97 (D88B6) XP® Rabbit mAb |
5169 |
WB IP IF F |
H M R Mk |
4 |
||
eIF4GI (D6A6) Rabbit mAb |
8701 |
WB IP IHC |
H M R |
2 |
||
eIF6 (D16E9) XP® Rabbit mAb |
3833 |
WB IP IF |
H M R Mk |
1 |
||
假尿书写器 |
Dyskerin (D6N4K) Rabbit mAb |
53234 |
WB |
H M Mk |
||
靶点 初筛 |
N6-mA Methyltransferase Antibody Sampler Kit |
69159 |
METTL3、METTL14、Virilizer、WTAP、Anti-rabbit IgG |
|||
Translational Control Antibody Sampler Kit |
9918 |
Phospho-Akt、Phospho-p70 S6 Kinase、Phospho-S6 Ribosomal Protein、Phospho-eIF2α、Phospho-4E-BP1、Phospho-eIF4E、Anti-rabbit IgG |
||||
Translation Initiation Complex Antibody Sampler Kit |
4763 |
eIF4A、eIF4A1、eIF4B、Phospho-eIF4B、eIF4E、Phospho-eIF4E、eIF4G、Phospho-eIF4G、eIF4H、Anti-rabbit IgG |
||||
其他RNA相关蛋白 |
LIN28A (D84C11) XP® Rabbit mAb |
3695 |
WB IF F |
H |
3 |
|
Nucleomethylin Antibody |
4789 |
WB |
H |
|||
3
有哪些研究套路可参考?
在m6A甲基化研究领域,芝加哥大学的何川教授对此贡献巨大,被公认为“RNA表观遗传学”这个全新研究领域的发起人之一。下文分享几篇10+分文章,供大家参考学习!
1.Nature cell biology(19分)| 低水平m6A mRNA甲基化通过影响AKT信号通路促进子宫内膜癌细胞的增殖和致瘤性
2018年8月,Nature cell biology 杂志发表了芝加哥大学的何川教授和Ernst Lengyel教授的最新研究成果。

(点击图片,查看原文)
本研究通过TCGA数据分析、甲基化水平检测、基于病毒载体在细胞和动物模型上进行基因表达水平操作(敲除、RNA干扰和过表达)。研究发现大约70%的子宫内膜肿瘤表现出m6A甲基化修饰的减少,这可能是由于甲基转移酶复合物组分METTL14突变或另一组分METTL3的表达降低。研究证实低水平m6A甲基化通过激活AKT信号途径增强了子宫内膜癌细胞的增殖和致瘤性。
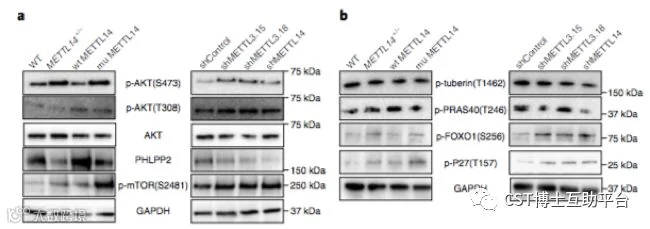
图7. 减少的m6A甲基化激活AKT(CST抗体的精彩表现)
总之,这些结果揭示了m6A mRNA甲基化为子宫内膜癌的致癌机制,并将m6A甲基化鉴定为AKT信号传导的调节因子。Working model见图8。
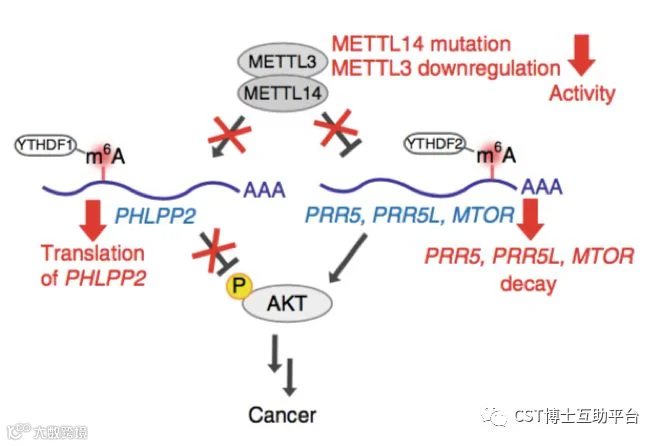
图8. 模型显示了通过降低m6A甲基化改变AKT信号以促进肿瘤进展
本文中所使用的CST抗体(Akt、mTOR信号通路)及其他相关产品见如下表。
产品名称 |
货号 |
应用 |
反应性 |
文献数 |
Phospho-mTOR (Ser2448) (49F9) Rabbit mAb (IHC Specific) |
2976 |
IHC |
H |
88 |
Phospho-Akt (Ser473) (D9E) XP® Rabbit mAb |
4060 |
WB IP IHC IF F |
H M R Hm Mk Dm Z B |
2449 |
Phospho-Akt (Thr308) Antibody |
9275S |
WB IP |
H M R |
652 |
Akt Antibody |
9272 |
WB IP IF F |
H M R Hm Mk C Dm B Dg Pg GP |
3828 |
Phospho-Tuberin/TSC2 (Thr1462) (5B12) Rabbit mAb |
3617T |
WB IP |
H M R |
39 |
Phospho-PRAS40 (Thr246) (C77D7) Rabbit mAb |
2997T |
WB IP IHC |
H M R Mk |
84 |
Phospho-FoxO1 (Ser256) Antibody |
9461T |
WB |
H M R Mk |
189 |
Rictor Antibody |
2140S |
WB |
H M R Mk |
38 |
Raptor (24C12) Rabbit mAb |
2280 |
WB IP |
H M R Mk |
166 |
2.Nature(41.57分)| m6A mRNA甲基化通过靶向IL-7/STAT5/SOCS信号通路控制T细胞的稳态
2017年8月,Nature杂志发表了来自暨南大学的尹芝南教授、耶鲁大学的Richard A. Flavell教授和Huabing Li教授的研究成果。

点击图片,查看原文
该研究通过构建m6A修饰酶METTL3的条件性敲除小鼠模型,系统性地研究了N6-腺苷甲基化修饰调控T细胞稳态的分子机制。研究发现,T细胞内敲除RNA N6-腺苷甲基化修饰酶后,小鼠的幼稚T(Naive T cells)细胞数量和比例显著上升,且该细胞无法进行正常的稳态增殖和分化。进一步研究发现N6-腺苷甲基化修饰可以通过靶向白介素7(Interleukin 7, IL-7)介导的细胞因子信号通路抑制分子 (Suppressor of cytokine signaling,SOCS) 信使RNA的快速降解,促进幼稚T细胞重编程(Reprogram)进而启动T细胞的增殖、分化。该研究使得开发靶向m6A的药物来缓解自身免疫疾病成为一种可能。Working model见图10。
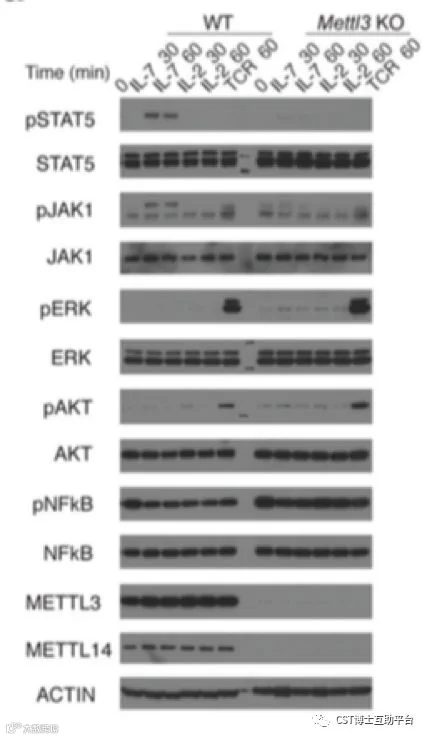
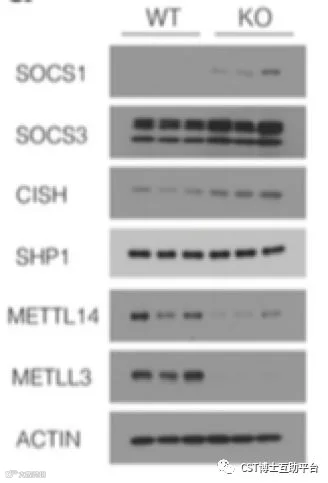
图9. Mettl3 敲除的幼稚T细胞中过表达的m6A靶基因Socs1,Socs3和Cish抑制了IL-7 / STAT5信号通路(CST抗体的精彩表现)
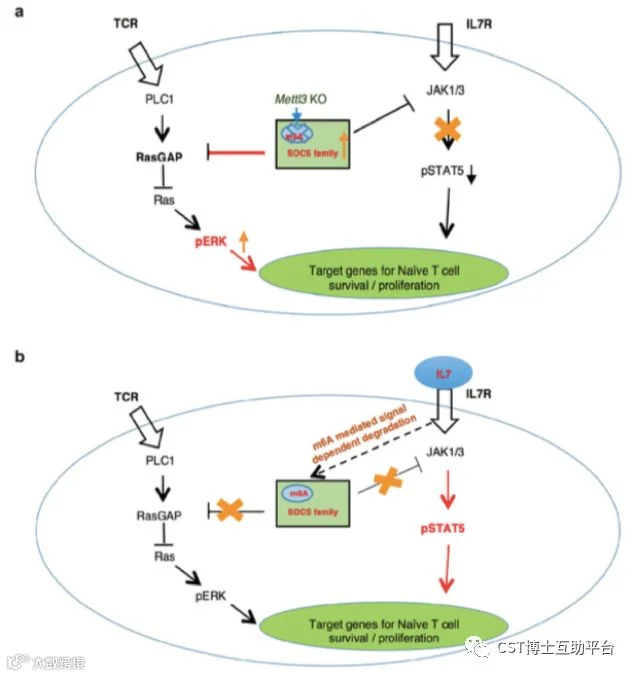
图11. m6A控制初始T细胞稳态的工作模型
本文中所使用的CST抗体及其他相关产品见如下表。CST提供一整套经典信号通路(Akt、Jak-Stat、MAPK、NF-KB、IL信号通路)抗体,m6A修饰酶抗体、CD Markers 、流式抗体、细胞因子产品等。
靶点 |
产品名称 |
货号 |
应用 |
反应性 |
文献数 |
Akt |
Akt (pan) (11E7) Rabbit mAb |
4685 |
WB IP IHC IF F |
H M R Mk |
489 |
Phospho-Akt (Ser473) (D9E) XP® Rabbit mAb |
4060 |
WB IP IHC IF F |
H M R Hm Mk Dm Z B |
2449 |
|
Stat5 |
Stat5 (D3N2B) Rabbit mAb |
25656 |
WB IP IHC IF |
H |
1 |
Stat5 (D2O6Y) Rabbit mAb |
94205 |
WB IP CHIP |
H M R |
4 |
|
Phospho-Stat5 (Tyr694) (D47E7) XP® Rabbit mAb |
4322 |
WB IF F |
H M |
50 |
|
Jak1 |
Jak1 (6G4) Rabbit mAb |
3344 |
WB IHC |
H M R |
28 |
Phospho-Jak1 (Tyr1034/1035) Antibody |
3331 |
WB |
H M |
60 |
|
ERK |
p44/42 MAPK (Erk1/2) Antibody |
9102 |
WB IP IHC |
H M R Hm Mk Mi Z B Pg Sc |
2903 |
Phospho-p44/42 MAPK (Erk1/2) (Thr202/Tyr204) Antibody |
9101 |
WB IP IF F |
H M R Hm Mk Mi Dm Z B Pg Ce |
3334 |
|
NF-κB p65 |
NF-κB p65 (D14E12) XP® Rabbit mAb |
8242 |
WB IP IHC IF F CHIP |
H M R Hm Mk Dg |
602 |
Phospho-NF-κB p65 (Ser536) (93H1) Rabbit mAb |
3033 |
WB IP IF F |
H M R Hm Mk Pg |
824 |
|
FTO |
SOCS1 (A156) Antibody |
3950 |
WB |
H M R Mk |
23 |
SOCS3 Antibody |
2923 |
WB |
H M R |
45 |
|
CISH |
CISH (D4D9) Rabbit mAb |
8731 |
WB IP IF F |
H M |
3 |
Annexin |
Annexin V Antibody |
8555 |
WB |
H M R Mk |
4 |
METTL3 |
METTL3 (D2I6O) Rabbit mAb |
96391 |
WB IP |
H M R Mk |
|
METTL14 |
METTL14 (D8K8W) Rabbit mAb |
51104 |
WB |
H M R Mk |
|
CD Markers |
CD3 (17A2) Rat mAb (violetFluor™ 450 Conjugate) |
38527 |
F |
M |
|
CD28 (CD28.2) Mouse mAb (PE-Cy7® Conjugate) |
82076 |
F |
H |
||
CD4 (RPA-T4) Mouse mAb (violetFluor™ 450 Conjugate) |
26755 |
F |
H |
||
CD8α (D8A8Y) Rabbit mAb |
85336 |
WB IP IHC IC |
H |
||
CD44 (156-3C11) Mouse mAb |
3570 |
WB IP IHC IF F |
H |
75 |
|
CD45 (D9M8I) XP® Rabbit mAb |
13917 |
WB IP IHC IF F |
H |
3 |
|
FoxP3 |
FoxP3 (D6O8R) Rabbit mAb |
12653 |
IHC IF F |
M |
3 |
IL-4 |
IL-4 (D19A10) Rabbit mAb |
12227 |
WB F |
H |
|
细胞因子 |
Human Interleukin-2 (hIL-2) |
8907 |
|||
Human Interleukin-4 (hIL-4) |
8919 |
||||
Mouse Interleukin-7 (mIL-7) |
5217 |
||||
3.Cancer Cell(22.84分) | FTO作为m6A RNA去甲基化酶在急性骨髓性白血病中发挥致癌作用
2017年1月,Cancer Cell杂志发表了来自芝加哥大学的Jianjun Chen教授的研究成果。

点击图片,查看原文
本研究表明,FTO在具有t(11q23)/ MLL重排,t(15; 17)/ PML-RARA,FLT3-ITD和/或NPM1突变的AML中高表达。FTO通过降低靶基因(例如ASB2和RARA)mRNA转录物中的 m6A水平,调节这些基因的表达,增强白血病癌基因介导的细胞转化和白血病发生,并抑制全反式维甲酸(ATRA)诱导的AML细胞分化。研究证明了 m6A甲基化和相应的蛋白质在癌症中的功能重要性,并为白血病发生和药物反应提供了新的思路。Working model见图13。
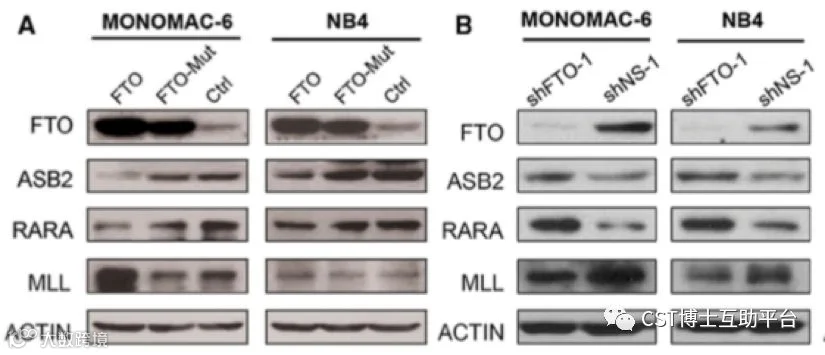
图12. ASB2和RARA是AML中FTO的两个重要的靶基因(CST抗体的精彩表现)
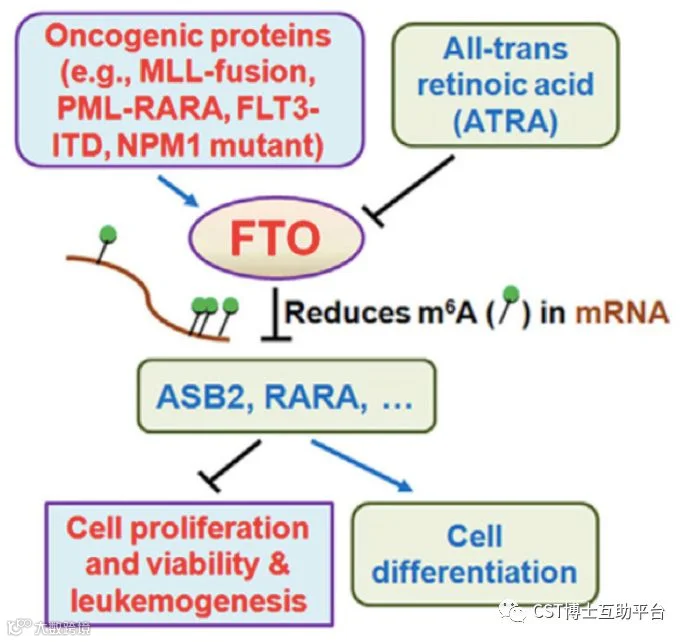
图13. FTO在白血病发生和ATRA诱导白血病细胞分化中的作用和潜在机制的示意图
本文中所使用的CST抗体及其他相关产品见如下表。
靶点 |
产品名称 |
货号 |
应用 |
反应性 |
文献数 |
FTO |
FTO (D2V1I) Rabbit mAb |
14386 |
WB |
H M R |
1 |
MLL1 |
MLL1 (D2M7U) Rabbit mAb (Amino-terminal Antigen) |
14689 |
WB IP |
H M R Mk |
2 |
Actin |
β-Actin (8H10D10) Mouse mAb |
3700 |
WB IHC IF F |
H M R Hm Mk Dg |
618 |
4. Cell Research(15.39分)|首次发现m6A修饰调控细胞自噬的重要功能
2018年7月,Cell Research杂志发表了来自中山大学的崔隽教授和任间教授课题组的研究成果。

点击图片,查看原文
该研究发现m6A去甲基化酶FTO能够去除自噬相关基因ULK1的3′-UTR上m6A修饰,抑制ULK1的降解,从而促进细胞自噬流的进程。研究人员发现FTO是细胞自噬的正调控因子,它可以正向调控ULK1的蛋白水平。结合最新的m6A-seq技术,研究人员发现ULK1的3′-UTR存在3个m6A化位点,随后证实FTO通过去除这3个腺苷酸上的甲基化修饰稳定ULK1的蛋白量。m6A修饰是ULK1发生降解的信号,在自噬发生过程中,FTO可以显著性地切除ULK1的3′-UTR的甲基化修饰,抑制“阅读器”YTHDF2对于ULK1的识别和降解,推动自噬发生(见如下图14)。
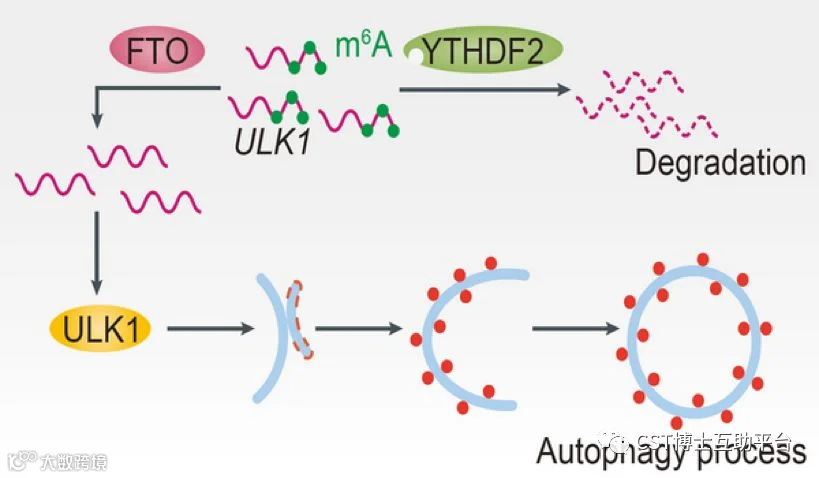
图14. 自噬启动时,FTO去除ULK1的甲基化修饰,抑制其YTHDF2依赖的降解,从而上调ULK1蛋白量,促进自噬过程。
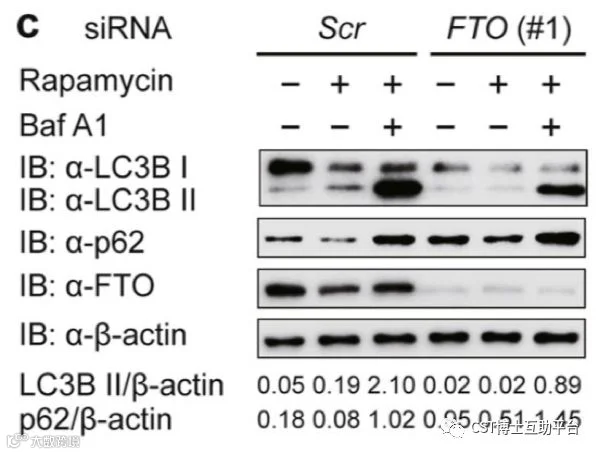
图15. FTO介导的ULK1转录物去甲基化通过上调ULK1蛋白丰度来促进自噬通量(CST抗体的精彩表现)
本文中所使用的CST抗体及其他相关产品见如下表。CST提供一整套相关信号通路(mTOR、自噬、凋亡、焦亡、坏死性凋亡等)抗体等产品。
靶点 |
产品名称 |
货号 |
应用 |
反应性 |
文献数 |
LC3A/B |
LC3B (D11) XP® Rabbit mAb |
3868 |
WB IP IHC IF F |
H M R |
358 |
LC3A/B (D3U4C) XP® Rabbit mAb |
12741 |
WB IHC IF F |
H M R |
135 |
|
ULK1 |
ULK1 (D8H5) Rabbit mAb |
8054 |
WB IP |
H M R Mk |
126 |
Phospho-ULK1 (Ser555) (D1H4) Rabbit mAb |
5869 |
WB IP |
H M |
93 |
|
SQSTM1 |
SQSTM1/p62 Antibody |
5114 |
WB |
H M R Mk |
297 |
SQSTM1/p62 (D5L7G) Mouse mAb |
88588 |
WB IP IHC IF |
H |
5 |
|
SQSTM1/p62 (D5E2) Rabbit mAb |
8025 |
WB IP |
H Mk |
82 |
|
Phospho-SQSTM1/p62 (Thr269/Ser272) Antibody |
13121 |
WB IP |
H M R |
4 |
|
Beclin-1 |
Beclin-1 (D40C5) Rabbit mAb |
3495 |
WB IP |
H M R Mk |
219 |
Phospho-Beclin-1 (Ser15) (D4B7R) Rabbit mAb |
84966 |
WB |
H M |
3 |
|
Atg7 |
Atg7 (D12B11) Rabbit mAb |
8558 |
WB IP |
H M R |
106 |
FTO |
FTO (D6Z8W) Rabbit mAb |
31687 |
WB IP |
H |
1 |
FTO (D2V1I) Rabbit mAb |
45980 |
WB |
H M R |
1 |
|
Sampler Kit |
Autophagy Antibody Sampler Kit |
4445 |
Beclin-1、LC3A/B、、Atg3、Atg5、Atg7、Atg12、Atg16L1、Anti-rabbit IgG |
||
本节内容部分摘自中山大学新闻网。
5.Nature(41.57)|SMAD2/SMAD3通过m6A修饰参与干细胞分化

点击图片,查看原文
TGF-β通路是干细胞多能性的调控者,主要通过SMAD2和SMD3两个效应蛋白介导。研究人员首次发现SMAD2/3能够激活RNA修饰的蛋白,从而使得特定的mRNA变得不稳定和快速地降解。研究发现,存在METLL3-METLL14-WTAP复合物的直接互作,在干细胞分化时SMAD2/SMAD3调控着下游靶基因RNA水平的m6A水平提高,包括Nanog,使得干细胞向特定方向分化。Model见如下图16。
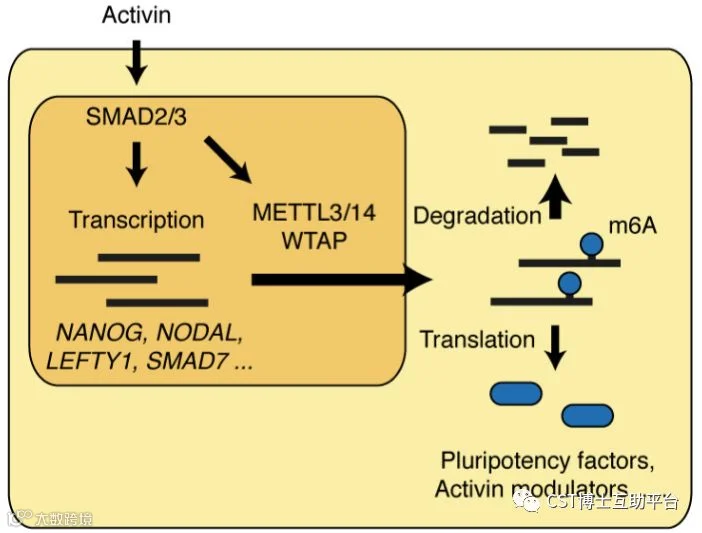
图16. 模型显示hPSC中激活素-NODAL信号传导和m6A沉积之间的相互作用
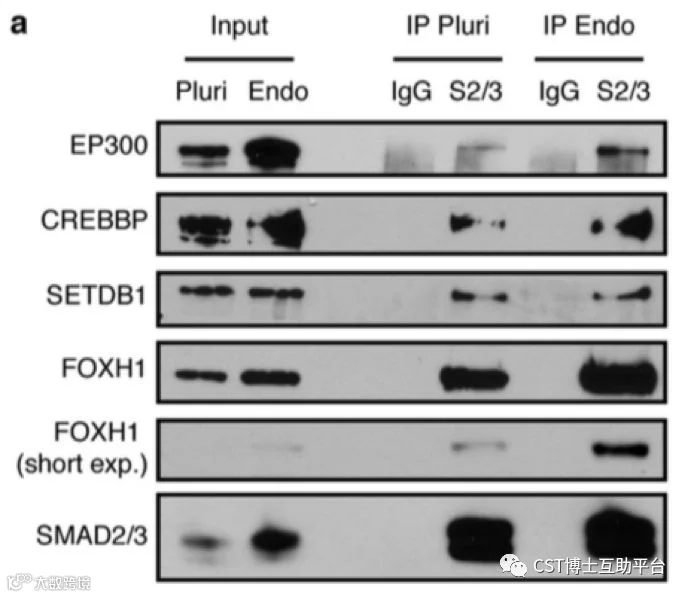
图17. hPSC中SMAD2 / 3转录和表观遗传辅助因子的功能特征(CST抗体的精彩表现)
本文中所使用的CST抗体及其他相关靶点产品见如下表。CST提供一整套发育相关信号通路(干细胞、ECS全能性和分化、Wnt、TGF-β、Hippo、Notch、Hedgehog信号通路等)抗体等研究工具。
产品名称 |
货号 |
应用 |
反应性 |
文献数 |
CBP (D6C5) Rabbit mAb |
7389 |
WB IP IF CHIP |
H M R Mk |
21 |
Nanog (D2A3) XP® Rabbit mAb (Mouse Specific) |
8822 |
WB IP IF F CHIP |
M |
19 |
Nanog (D73G4) XP® Rabbit mAb |
4903 |
WB IHC IF F |
H |
66 |
Oct-4 Antibody |
2750 |
WB IHC IF F CHIP |
H |
67 |
Oct-4 (D7O5Z) Mouse mAb |
75463 |
WB IF F |
H M |
|
ESET (C1C12) Rabbit mAb(SETDB1) |
2196 |
WB IP IF |
H Mk |
4 |
Smad2/3 (D7G7) XP® Rabbit mAb |
8685 |
WB IP IF F CHIP |
H M R Mk |
88 |
Smad2/3 (D7G7) XP® Rabbit mAb (Biotinylated) |
12470 |
WB |
H M R Mk |
1 |
Smad1 (D59D7) XP® Rabbit mAb |
6944 |
WB IP CHIP |
H M Mk |
51 |
Phospho-Smad1 (Ser463/465)/ Smad5 (Ser463/465)/ Smad9 (Ser465/467) (D5B10) Rabbit mAb |
13820 |
WB IP IF F |
H M R |
36 |
Sox1 Antibody |
4194 |
WB IF |
M R |
16 |
Sox17 (D1T8M) Rabbit mAb |
81778 |
WB IF |
H |
2 |
Sox2 Antibody |
2748 |
WB IP CHIP |
H M |
63 |
Phospho-Sox2 (Ser250/Ser251) (A2I7G) Rabbit mAb |
92186 |
WB IP |
H M |
1 |
SnoN Antibody |
4973 |
WB IF |
H |
3 |
最后,简单总结下RNA甲基化研究的思路和相关实验方法、研究工具等,具体大家可参看上述的高分文章和中标的国自然标书。
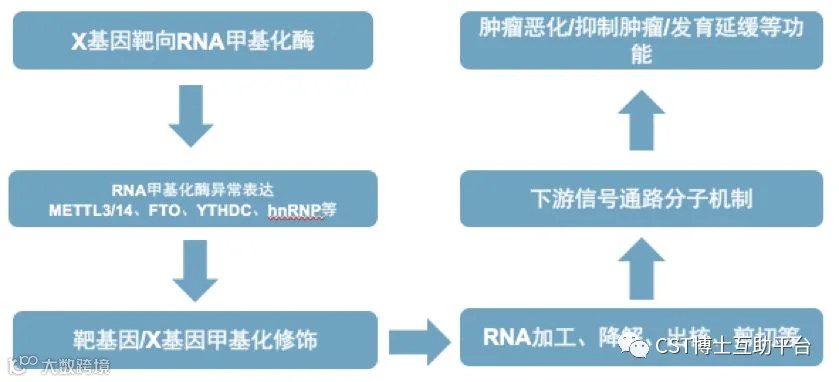
图16. RNA甲基化修饰基本研究思路
实验流程 |
研究工具或方案 |
检测RNA整体m6A水平 |
m6A RNA甲基化定量Kit |
定量PCR/MeRIP-qPCR |
qPCR商品化试剂盒、m6A抗体 |
MeRIP-Seq |
测序 |
基因敲除、敲低或过表达 |
RNA甲基化相关酶和下游靶基因的干扰or敲除or过表达的质粒、病毒、稳定细胞株、模式动物等 |
细胞/动物表型实验 |
细胞增殖、迁移、凋亡、自噬、周期、焦亡、衰老等; |
分子机制研究 |
基因表达分析:qPCR、WB、ELISA、IF、Flow、IHC、组织染色; |
蛋白-基因互作:Co-IP、ChIP、BioID、RNA pulldown、GST pulldown、EMSA等 |
|
下游靶基因研究:基因芯片、蛋白芯片、PTMScan蛋白质翻译后修饰高通量筛选、CRISPR/Cas9、组织芯片、信号通路生物信息分析; |
|
临床意义 |
继续深入研究上述实验挖掘到的甲基化酶或是被调控的下游通路基因的临床意义; |
基因有限公司作为Cell Signaling Technology (CST) 优质一级代理商,为您实验提供更优的支持和帮助。想要了解更多,请在本文后留言或联系您身边的基因有限公司员工。
