
生物化学家 Christian de Duve 于 1963 年引入的术语“自噬”源自希腊语,意思是“自食”,是细胞质成分降解和回收的基本过程。自噬的广义术语已分为不同类型,包括:1) 巨自噬:细胞内容物和细胞器被双膜自噬体吞噬的主要形式,该自噬体与溶酶体融合以降解,2) 微自噬:溶酶体直接与细胞质货物融合, 3) 伴侣蛋白介导的自噬:细胞质伴侣蛋白协助将靶标带到溶酶体的过程。
巨自噬,通常简称为自噬,其特征最具代表性。它受营养条件和细胞应激的高度调节,并在正常生理稳态中发挥重要作用。它经常在病理状况中受到影响,比如癌症、代谢紊乱、炎症性疾病、宿主防御和神经变性。
2016 年,诺贝尔生理学或医学奖授予 Yoshinori Ohsumi,以表彰其对酵母中自噬所需基因的初步鉴定。在酵母中发现的基因为接下来发现的在自噬中起关键作用的 20 多个自噬相关基因 (ATG) 铺平了道路。自噬通路的一般特征包括 1) 启动自噬,将货物隔离到吞噬细胞;2)自噬体延伸并成熟,成为全封闭结构;3)自噬体与溶酶体的融合。
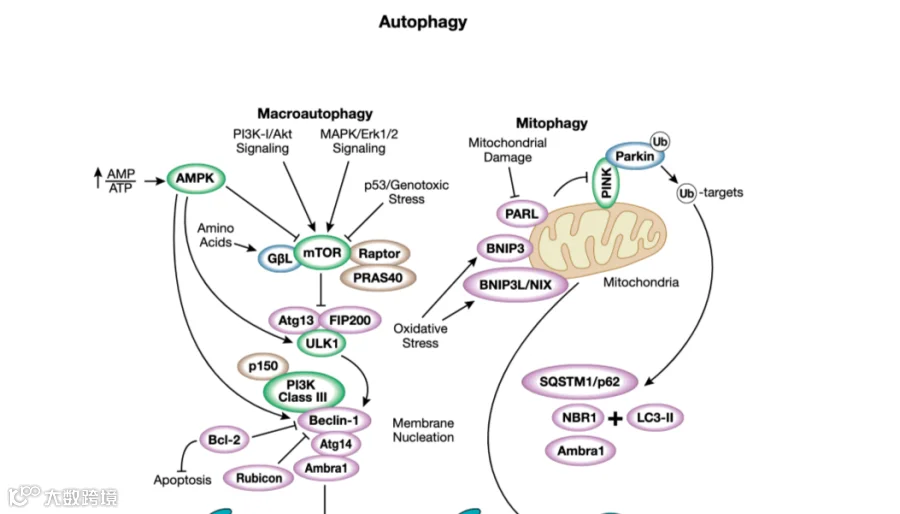
自噬的启动通常由细胞能量和营养传感器 AMPK 和 mTORC1 控制。这些主导性激酶在自噬的启动中具有此强彼弱的关系。这些激酶的靶标之一是自噬激酶 ULK1。AMPK 将 ULK1 的多个位点(包括 Ser317 和 Ser555)磷酸化并激活UKL1;而 mTORC1 将 ULK1 的Ser757 位点磷酸化并抑制其活性。自噬体的成熟需要激活 ULK1 和多蛋白复合物的募集,这样可以激活脂质激酶 III 类磷脂酰肌醇 3 激酶 (VSP34)。自噬体成熟所需的 ULK1 底物包括 ATG13、Beclin-1、ATG14 和 VSP34。自噬体成熟还需要两个以渐进方式起作用的泛素样结合系统。
在第一步中,ATG12 与 ATG5 结合。随后是磷脂酰乙醇胺 (PE) 与 LC3 或 GABARAP 家族成员的脂质缀合。第二步的结合过程,通常称为 II 型形式,允许掺入到自噬体膜中,并且经常用作自噬的标记物。最后,自噬体的内容物被溶酶体内的 pH 敏感水解酶降解。抑制溶酶体酸化的药物(如氯喹和巴弗洛霉素 A1)常用于抑制自噬的后期。
自噬不仅仅是细胞内成分的大量降解。相反,它也可以是一个高度选择性的过程,旨在降解特定的细胞器、病原体和蛋白质。线粒体 (mitophagy)、ER (reticulophagy 或 ER-phagy)、核糖体 (ribophagy)、过氧化物酶体 (pexophagy)、溶酶体 (lysophagy) 和细胞核 (nucleophagy) 以及蛋白质聚集体 (aggrephagy)、脂滴 (lipophagy) 和细胞内病原体 (xenophagy)的特定细胞器降解通路已被描述。
自噬还可以靶向选择性蛋白质(如铁蛋白)在调节铁代谢中很重要,并通过铁死亡控制铁依赖性细胞死亡。细胞成分经常被货物受体靶向自噬体(例如 SQSTM1/p62 家族中的那些),它们通过与 LC3 家族成员的相互作用将其靶向内容物与自噬体结合。选择性自噬通常与越来越多的靶向选择成分的特定货物受体有关。
在过去十年中,我们对健康和疾病背景下的非选择性和选择性自噬的理解取得了巨大进步。许多研究已经描述了自噬对癌症的积极和消极作用。虽然通过自噬清除损伤细胞器可以防止肿瘤发生,但癌细胞通常具有更高的自噬率,这是在营养贫乏的环境中生存所必需的。抑制自噬,无论是通过基因敲除关键自噬基因,还是在药理学上使用氯喹及其衍生物等药物,都可以减少肿瘤负担。
最近,特定的自噬抑制剂(例如抑制 ULK1 的化合物)已经提供了新的治疗机会。相反,诱导自噬(例如通过抑制 mTORC1)也可能证明对其他疾病具有临床影响。病原体通过异体自噬而发生的选择性自噬是宿主防御的一个重要因素。微生物毒力可能会受到它们破坏自噬防御机制的能力的影响。像激酶 TBK1 这样的蛋白质在先天免疫通路和增强自噬中发挥双重作用。自噬的保护作用也是神经退行性疾病的共同特征。通过线粒体自噬选择性降解受损线粒体已被认为是能量稳态的重要因素,并且在神经变性中经常被破坏。参与线粒体自噬的蛋白质(如 PINK1 和 Parkin)可能会在帕金森病和其他疾病中发生突变。在该通路中,泛素在 Ser65 位点的 PINK1 磷酸化导致泛素 E3 连接酶 Parkin 的激活和募集到线粒体,这导致线粒体靶标的泛素化,从而被自噬货物受体识别。正在研究鉴定线粒体自噬激活剂以治疗这些疾病。
Cell Signaling Technology TM(CST® )开发了经过充分验证的抗体和试剂盒,旨在满足自噬研究人员的需求并推动治疗发展。可以使用我们高引用的针对 LC3 家族成员、货物受体和关键自噬激酶(包括 AMPK、mTORC1、TBK1、PINK1 和 ULK1)的磷酸化特异性靶标的抗体来分析自噬。请访问 www.cellsignal.cn 以了解更多详情。
参考文献:
Yang Y, Klionsky DJ. Autophagy and disease: unanswered questions. Cell Death Differ. 2020 Mar;27(3):858-871. doi: 10.1038/s41418-019-0480-9. Epub 2020 Jan 3. PMID: 31900427; PMCID: PMC7206137.
Mizushima N, Levine B. Autophagy in Human Diseases. N Engl J Med. 2020 Oct 15;383(16):1564-1576. doi: 10.1056/NEJMra2022774. PMID: 33053285.
Miller DR, Thorburn A. Autophagy and organelle homeostasis in cancer. Dev Cell. 2021 Apr 5;56(7):906-918. doi: 10.1016/j.devcel.2021.02.010. Epub 2021 Mar 8. PMID: 33689692; PMCID: PMC8026727.
Giorgi C, Bouhamida E, Danese A, Previati M, Pinton P, Patergnani S. Relevance of Autophagy and Mitophagy Dynamics and Markers in Neurodegenerative Diseases. Biomedicines. 2021 Feb 4;9(2):149. doi: 10.3390/biomedicines9020149. PMID: 33557057; PMCID: PMC7913851.
Poillet-Perez L, White E. Role of tumor and host autophagy in cancer metabolism. Genes Dev. 2019 Jun 1;33(11-12):610-619. doi: 10.1101/gad.325514.119. PMID: 31160394; PMCID: PMC6546058.
Deretic V. Autophagy in inflammation, infection, and immunometabolism. Immunity. 2021 Mar 9;54(3):437-453. doi: 10.1016/j.immuni.2021.01.018. PMID: 33691134; PMCID: PMC8026106.
Egan DF, Chun MG, Vamos M, Zou H, Rong J, Miller CJ, Lou HJ, Raveendra-Panickar D, Yang CC, Sheffler DJ, Teriete P, Asara JM, Turk BE, Cosford ND, Shaw RJ. Small Molecule Inhibition of the Autophagy Kinase ULK1 and Identification of ULK1 Substrates. Mol Cell. 2015 Jul 16;59(2):285-97. doi: 10.1016/j.molcel.2015.05.031. Epub 2015 Jun 25. PMID: 26118643; PMCID: PMC4530630.
联系CST产品专员,获取电子版/纸质版资料

基因有限公司作为 Cell Signaling Technology (CST)优质一级代理商,为您的实验提供更优的支持和帮助。想要更多相关知识和资料,或了解促销详情请在本文后留言或联系您身边的基因有限公司员工。
