



本文原文刊于
《今日声学》(Acoustics Today) 17卷 1期
标题:
Ultrasonic Hearing in Cats and Other Terrestrial Mammals
作者:
M. Charlotte Kruger, Carina J. Sabourin, Alexandra T. Levine, and Stephen G. Lomber
主要内容包括:4种陆栖哺乳动物可感知到的超声波频段,超声感知能力的生物学重要性,以及目前已知的动物处理超声波时所使用的脑部分区。

当你看到“超声(ultrasound)”这个词,首先进入你脑海的是什么?
大部分人可能会联想到超声孕检、航海及航空探测等领域,也有人会想到蝙蝠、海豚、鲸鱼这些众所周知利用超声波进行回声定位的动物。那么,你会联想到和人类一样生活在陆地上的其他哺乳动物吗?
事实上,对于不能飞的陆地哺乳动物的超声感知能力,我们的了解程度确实比较有限。

我们关于超声波的了解

超声波之所以被认为不同于“普通”声波,是因为它频率高,人类听不见。人类的听力上限约为20kHz,因而频率超过20kHz的声波就是超声波了,这是超声波定义中公认的部分。显然,这个标准是根据人类的听觉感知范围制定出来的,较为主观,本质上并没有生物学基础。
人类尽管无法听到超声波,却可以在日常生活中对它的存在加以利用,其中我们最熟悉的莫过于超声波的临床应用(Ketterling and Silverman, 2017),包括孕期扫描、病理观察、碎肾结石(Simon et al., 2017)等等。在工业上,超声波还被用于物品厚度和质量的无损检测。虽然超声波应用场景广泛,但暴露于机载超声中也会导致人体产生恶心、头晕、注意力无法集中等不良反应(Leighton et al., 2020)。然而对于动物来说并不是这样的,出于各种原因,在人类开始利用超声频段之前很久,动物就已经在使用超声波了。

图1. 声压级为60dB时几种动物的听觉频率范围。小家鼠和棉鼠(Mus musculus and Sigmodon Hispidu)(Masterson and Heffner, 1980),亚洲象(Elephas maximus)(Heffner and Heffner, 1982),家猫(Felis catus)(Heffner and Heffner, 1985),家狗(Canis lupus familiaris)(Heffner, 1983),昭短尾叶鼻蝠(Carollia perspicillata)(Koay et al., 2003),以及通常人类应用的声波频率范围(Jensen, 2007; Carovac et al., 2011; Harvey et al., 2014)

听觉通路与超声波
动物探测和感知到很大频率范围和强度范围内声音的能力来自于它们的听力系统。声波穿过外耳和中耳,最后在内耳被传导到耳蜗上。耳蜗将不同频率和强度的声音转换成可以被脑部解读的电信号。这些电信号沿着听觉通路,从耳蜗开始,经过脑干,最终被丘脑中的核传递到它们的终点——听觉皮层。

听力频率范围测量

我们用听力测定实验来考察不同物种的超声感知能力。声音能被探测到的临界点被称为可听阈值(audibility threshold)。精神物理学方法常常被用来测量非人类物种的感知阈值——通过给测试对象施加一个物理刺激,来测量由此引起的神经和/或知觉响应。心理声学作为神经心理学的一个分支,内容涉及用各种条件方法来分析听觉刺激和神经活动之间的关系(Dent,2017)。不同条件下的测试结果通常用听力图来展示。(图2)

啮齿类动物
许多小型啮齿类动物通过超声波进行求偶、交配、繁殖、争夺领地、警示同类等交流活动(Arch and Narins, 2008)。雄性小鼠接收到雌性信息素,就会发出48kHz~79kHz的超声波,吸引对方将它们当作潜在伴侣(Gourbal et al., 2004),这显示了小鼠间的超声交流有一定的性别特异性。另外,在交配时雄鼠会发出高达75kHz的叫声(Matsumoto and Okanoya, 2016),而幼鼠如果被从窝里取出,也会发出50kHz~80kHz的离穴呼叫(Hofer et al., 2001)。小鼠可以在60dB的声压水平下听到1.5kHz~92kHz频段的声音(Hofer et al., 2001),因而幼鼠的呼叫确实能引起母亲的搜索(Dunlap and Liu,2018)
与小鼠相似,成年大鼠发出超声波主要是有两个交流目的:以22kHz声音警告同类出现危险和以50kHz声音进行社交合作和友好行为(Wright et al., 2010)。大鼠的听力范围在250Hz~80kHz之间,幼鼠的离穴呼叫频段为40kHz~65kHz,当听到雄鼠50kHz的叫声时,雌鼠会表现出一系列吸引行为来增加交配的可能性(Portfors, 2007)。综上,啮齿类动物的生存离不开超声感知。
食肉动物
不同于啮齿类动物,有关食肉动物的超声听力演化及生物学重要性的数据非常少。除了食肉啮齿动物(如北方蝗虫Onychomys leucogaster)以外,我们已知的食肉动物很少使用超声频率进行通讯交流(Brown et al. 1978; Farley et al., 1987)。不过,许多食肉动物都可以感知到超声频率。这或许是因为,在历史上的某一时期,食肉动物的共同祖先曾用超声波侦察过猎物(Heffner and Heffner, 1985; Kelly et al., 1986)。不过,就像上文介绍的那样,鼠类这样的猎物主要以超出食肉动物听力上限的频率进行交流(Kelly and Masterton, 1977)。
Phillips和他的同事(1988)认定,雪貂(Mustela putorius furo)可以听到40Hz~40kHz的声音。相较于其他哺乳动物,雪貂的听力出现得比较晚,因而它们是用于研究听觉皮层的发育、组构和可塑性的绝佳选择(Moore, 1982)。在新生雪貂的耳道开放之前,它们通常会发出高于16kHz的声音,和鼠类一样,处于哺乳期的雌性雪貂会对幼崽的高频叫声有所反应(Shimbo, 1992)。此外,雪貂还和人类的听力范围有很大重叠。对于有关听力及听力损失的各方面研究而言,雪貂是一个很有帮助的模型。
另一种常用作听觉研究模型的食肉动物是家猫(felis catus)。我们通常认为,猫的听力敏感频段为5kHz~32kHz。尽管各类文献中记述的猫的听力频率范围存在显著差异(图3),但猫可以听到超声波这一点是被广泛认同的。一般资料里记录猫的听力下限大约为125Hz,而上限还不确定。
大多数资料里记录的猫的听力上限实际是测试的最高频率,这个上限值通常不会大于60kHz(图3),也有些时候,上限值会被写成相关领域研究中使用过的最高测试频率,将电脉冲施加于听觉通路神经元的电模拟实验和行为实验即是如此。不过有一个例外,Heffner和Heffner(1985)的测试频率达到92kHz,他们得到猫的听力上限为85kHz。由此,猫的听力上限可能超过60kHz,并且它们大脑皮层中可能存在专门处理超声频率的神经元。


超声频率的皮层表征
小鼠
Tsukano和他的同事(2015)认为,背内侧区(DM)从前被归为A1背区的一部分,但实际它是专用于感知超声波的一块独立区域。该分区内存在对发声产生高度响应的神经元,响应频率高达40kHz——揭示了小鼠脑皮层中的某些神经元是如何对与日常行为相关的声音特征做出最佳响应的。这样的皮层组构被发现也存在于其他依赖超声生存的啮齿类动物脑中。
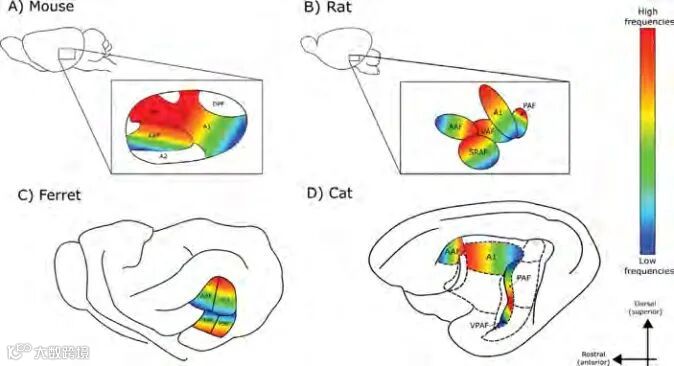
图4. 四种哺乳动物听觉皮层的音频拓扑组构。A)小鼠;B)大鼠;C)雪貂;D)猫。脑部矢状面示意中划分了:初级听皮层Ⅰ区(A1),次级听皮层Ⅱ区(A2),听皮层前区(AAF),背后区(DPF),超声区(UF),听皮层后区(PAF),腹侧区(VAF),鼻上区(SRAF),伪大脑侧裂后区(PPF),大脑侧裂上后区(PSF)和腹侧后区(VPAF)。
大鼠
和小鼠不同的是,大鼠皮层中并没划分出超声区,不过有研究人员(Kalatsky et al., 2005)认为这个区域很可能是存在的,因为大鼠和小鼠脑中的音频拓扑组构极为相似。总之,尽管已经发现了啮齿类动物皮层组构的共同之处,我们仍需进一步加深对它们听觉皮层中超声相关区域的研究,也许就会得到可以拓展至其他哺乳动物的普适性发现。
雪貂
我们认为对于小鼠、大鼠以及大多数其他哺乳动物,听觉皮层各分区上的特征频率都是从高到低头尾向分布的,并且在相邻分区的边界处发生反转(Bizley et al., 2005)。然而,这种反转并不存在于雪貂脑皮层中。对于雪貂,A1和AAF的频率梯度在背侧相遇,并向腹侧降低(图4C)(Kaas, 2011),因而A1和AAF的音频拓扑组构都是越接近背尖频率越高的。雪貂A1的生理特性(如音频拓扑组构和神经元特性)和猫的A1相似(Kaas,2011),但从听力图可以看出,雪貂的特征响应频率整体低于猫(图2)。
猫
Hall和Lomber(2015)确定了猫的听觉皮层上有A1、AAF、PAF和VPAF四个功能不同的拓扑组构并论述了相邻区域间的音频拓扑梯度反转(图4D)。随着A1从大脑侧裂后外侧沟(PES)的前部延伸至大脑侧裂前外侧沟(AES)的后部,其拓扑梯度增加。在PES的后边缘,A1达到其拓扑梯度的最小值,并在接近PAF时形成一个低频反转边界(Hall and Lomber,2015)。高频反转边界存在于A1-AAF和PAF-VPAF边界,这里很可能发现具有超声选择性的神经元。



但是,高频反转边界也可能包含宽频调谐的神经元亚群,用于整合A1和AAF上较宽频带的超声感知输入。研究人员有时会避免探测边界附近的这些区域,以防意外记录邻近区域的神经元而造成“污染”(Carrasco et al., 2015)。也正因为采取了这种措施,研究可能无法记录到真正具有最高特征频率的神经元,从而对我们理解猫和其他陆生哺乳动物听觉皮层接收超声刺激后的表现造成了限制。

结语

超声对许多动物的生存至关重要,这一点由某些物种脑部专用于超声响应的皮层空间大小就能证明。尽管我们知道超声波具有一定的生物学重要性,但许多陆生哺乳动物的超声听力可能一直被我们低估了,像文中讨论的一些动物,它们的超声听力范围至今没能被最终确定,它们脑中用于处理超声的确切区域也依旧难以定论。我们期待进一步的研究可以明确超声频率在整个听觉神经科学中的角色和定位。
译文仅供学习交流

如有不妥,欢迎指正
原文地址:https://acousticstoday.org/ultrasonic-hearing-in-cats-and-other-terrestrial-mammals-m-charlotte-kruger/

免责声明:本文旨在传递更多科研资讯及分享,所有其他媒、网来源均注明出处,如涉及版权问题,请作者第一时间后台联系,我们将协调进行处理(按照法规支付稿费或立即删除),所有来稿文责自负,两江仅作分享平台。转载请注明出处,如原创内容转载需授权,请联系下方微信号。
