

蔷薇类植物(rosids)是被子植物中物种最丰富的类群之一,包含17个目、135个科,约7万个物种,占被子植物总数的四分之一以上。该类群形态与生态类型多样,涵盖草本、灌木、乔木、藤本、寄生及多肉植物等,广泛分布于温带与热带森林。蔷薇类不仅在生态系统中占据重要地位,还具有极高的经济价值,包括蔷薇科水果、十字花科蔬菜、豆科作物以及锦葵科、壳斗科等主要木本植物类群,与人类生活密切相关。
揭示蔷薇类植物的演化历史对理解被子植物的演化机制至关重要。然而,其内部系统关系长期存在争议。基于叶绿体基因的研究将蔷薇类划分为Fabids和Malvids两大分支,但在核基因系统发育分析中,这两大类群并不构成单系。此外,蔷薇类的姐妹群也未达成共识,APG IV系统将其与葡萄目(Vitales)归为一类,但部分研究则支持虎耳草目(Saxifragales)为其近缘类群。这些系统发育冲突限制了对蔷薇类整体演化规律的认知。
高分辨率核基因组研究解析蔷薇类系统发育
近日,内蒙古大学/复旦大学黄建勋研究员与美国宾夕法尼亚州立大学马红教授团队在《JIPB》发表题为“Nuclear phylogenomics provide evidence to clarify key morphological evolution and whole genome duplication across rosids”的研究论文。研究整合203个新测转录组和2个浅层基因组数据,联合公共数据库共纳入316个蔷薇类物种,覆盖全部16个目和83个科,利用七组低拷贝核基因重建了迄今最全面、分辨率最高的蔷薇类核基因系统发育树。
研究有效澄清了Geraniales、Crossosomatales与Myrtales等类群间的深层系统发育关系,并揭示了多个核-质体系统发育冲突,可能源于远缘杂交、基因流或不完全谱系排序(ILS)。通过基因树调和方法,团队在20个科中检测到27次全基因组复制(WGD)事件,其中18次为首次报道或对已有事件的定位修正。
全基因组复制推动适应性辐射
多数WGD事件集中发生在白垩纪-古近纪交界(K-Pg界线)及古新世至始新世阶段,恰逢重大地质气候剧变期。K-Pg灭绝事件导致大量生态位空缺,可能为经历多倍化的蔷薇类植物提供了演化优势。WGD通过基因剂量效应增强植物抗逆性与生长稳健性,同时基因冗余为新功能演化提供空间,促进性状创新与适应性辐射。
统计分析显示,在17个发生WGD的家族级以上类群中,其物种丰富度显著高于姊妹类群(平均1,698.2 vs. 914.6,P = 0.046),支持WGD对多样化的促进作用。研究还发现,WGD与多个关键形态创新相关,如十字花科花部简化与干果结构、葫芦科卷须演化、蔷薇科苹果亚族子房结构转变等,表明基因组复制在关键分类单元形成中发挥重要作用。
形态演化的简约化趋势
通过对18组关键形态性状(花器官、叶序、果实类型等)进行祖先状态重建,研究揭示了蔷薇类向形态简约化演化的总体趋势,表现为花瓣减少、雄蕊数目简化、子房由下位转向上位等特征,在十字花目、壳斗目、蔷薇目和葫芦目中尤为明显。
协同演化驱动生态成功
研究指出,蔷薇类在白垩纪的迅速崛起是地质、气候与生物因素协同作用的结果。板块运动与温暖气候为扩张提供了有利环境,火山活动与海平面变化增加了生态复杂性。传粉昆虫(如蜜蜂、鳞翅目)的兴起推动了繁殖策略演化,而植食性昆虫(如甲虫、象甲)和早期哺乳动物、鸟类的取食行为则可能促进了防御机制与种子传播方式的演化。
进入古近纪后,K-Pg灭绝事件、古新世-始新世极热事件(PETM)及后续降温过程对植物生存构成严峻挑战。蔷薇类凭借WGD带来的基因组可塑性与适应潜力,在剧烈环境变动中实现快速多样化。
总结与展望
蔷薇类植物的成功并非偶然,而是基因组复制、形态创新与生物互作共同驱动的结果。该研究从系统发育、基因组演化与形态重建多维度描绘了蔷薇类的演化蓝图,为理解植物大类群在全球生态系统中的崛起提供了关键证据。未来深入探究其应对古环境变化的机制,有助于评估植物在现代气候变化下的适应潜力,并为预测长期演化趋势提供参考。蔷薇类的演化策略或可成为应对当前气候挑战的“植物应变模型”。
本研究由赵义勇、余迪、郭文钰、黄洁、郭婧、孙苗、胡宜、Douglas E. Soltis、Pamela S. Soltis、马红和黄建勋等共同完成。赵义勇为论文第一作者,马红与黄建勋为共同通讯作者。研究得到国家自然科学基金、教育部重点实验室及国际合作项目支持。
Sun, M., Folk, R.A., Gitzendanner, M.A., Soltis, P.S., Chen, Z., Soltis, D.E., Guralnick, R.P. (2020). Recent accelerated diversification in rosids occurred outside the tropics. Nat. Commun. 11: 3333.
Wang, H., Moore, M.J., Soltis, P.S., Bell, C.D., Brockington, S.F., Alexandre, R., Davis, C.C., Latvis, M., Manchester, S.R., and Soltis, D.E. (2009). Rosid radiation and the rapid rise of angiosperm-dominated forests. Proc. Natl. Acad. Sci. U.S.A. 106: 3853–3858.
Leebens-Mack, J.H., Barker, M.S., Carpenter, E.J., Deyholos, M.K., Gitzendanner, M.A., Graham, S.W., Grosse, I., Li, Z., Melkonian, M., Mirarab, S., et al. (2019). One thousand plant transcriptomes and the phylogenomics of green plants. Nature 574: 679–685.
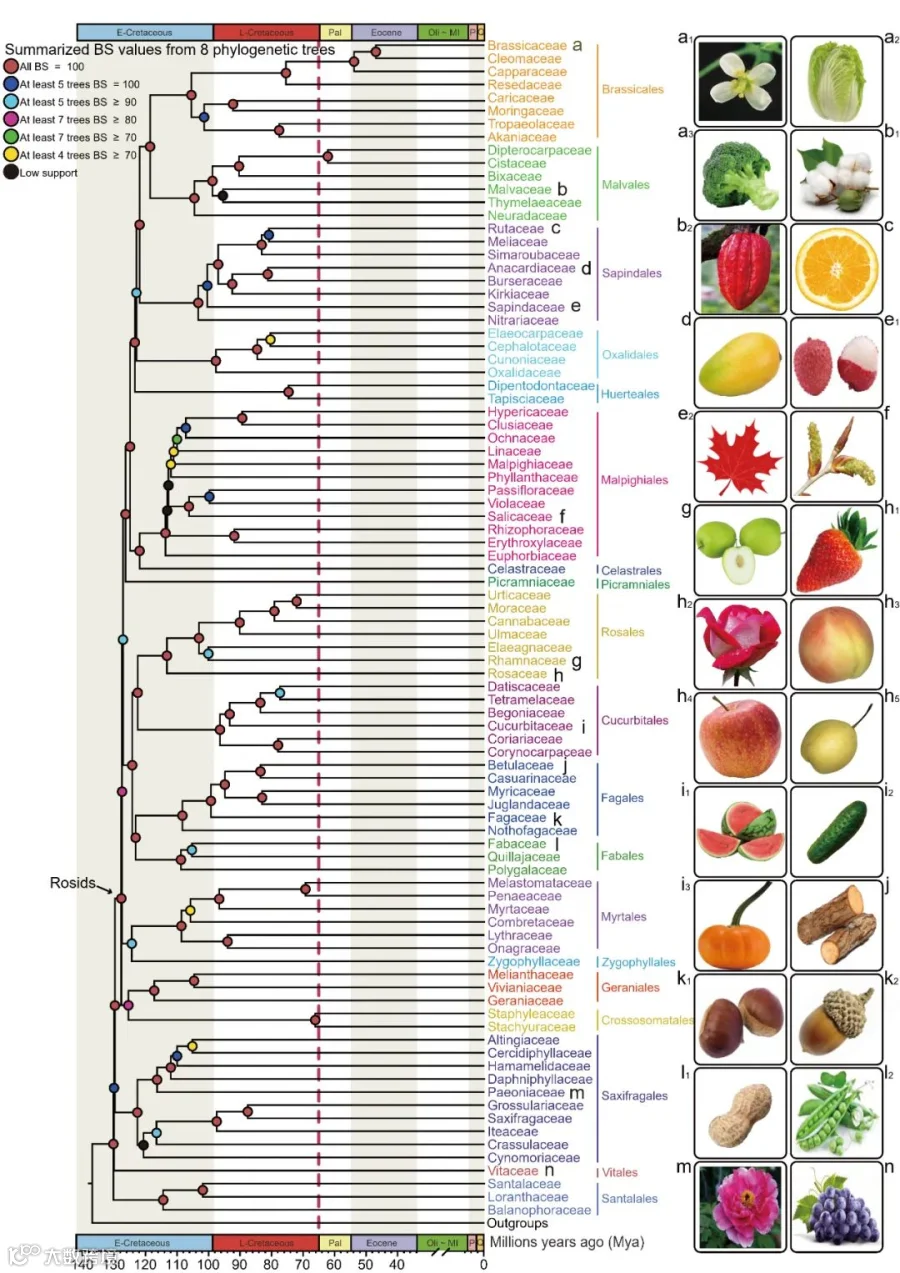
图1. 蔷薇类植物科之间的系统发育关系简图
本图展示了蔷薇类各科间的系统发育关系及分化时间(百万年前,Mya)。圆圈颜色表示来自八棵核基因树的支持率(bootstrap support, BS)类型;红色虚线标示K-Pg界线;末端名称按所属目着色。右侧为各目代表物种照片:a1拟南芥、a2卷心菜、a3西兰花、b1棉花、b2可可、c柠檬、d芒果、e1荔枝、e2枫树、f杨树、g枣树、h1草莓、h2玫瑰、h3桃、h4苹果、h5梨、i1西瓜、i2黄瓜、i3南瓜、j木材植物、k1板栗、k2橡实、l1花生、p豌豆、m牡丹、n葡萄。
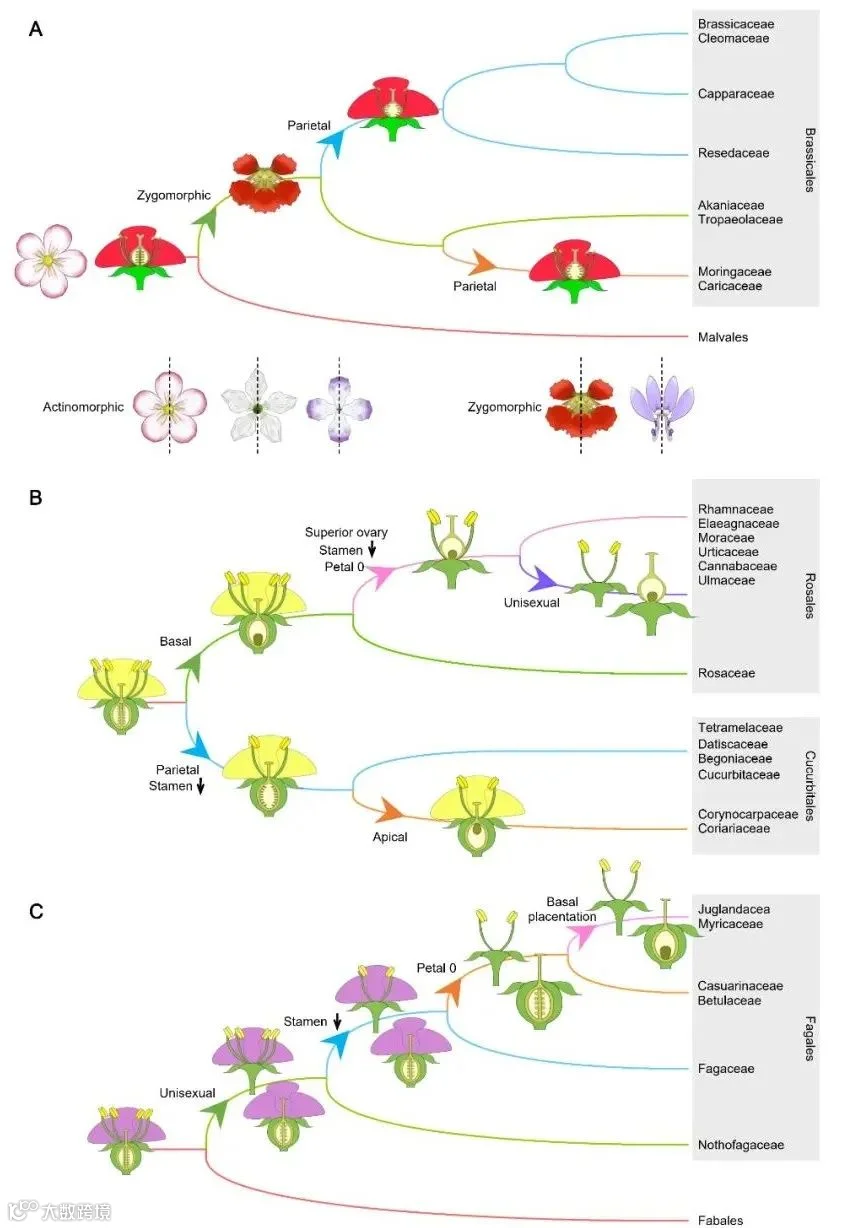
图2. 三个主要类群的祖先花部形态演化
展示十字花目+锦葵目(A)、蔷薇目+葫芦目(B)及壳斗目(C)的花部形态演化过程。系统发育树为简化版本,相同颜色分支表示具有相同祖先性状类型。